









Lipidy na rozhraní světů
| 28. 2. 2022„Dobré ploty dělají dobré sousedy“
Robert Frost, Mending Wall (1914)
Buněčná membrána buňku odděluje od okolí a chrání ji před vnějšími vlivy. Je-li porušena, buňka umírá. Můžeme ji tedy přirovnat ke skafandru astronautů. Ale příroda došla ve vývoji mnohem dál než lidstvo a s vlastnostmi buněčné membrány se naše skafandry nemohou ani zdaleka rovnat. Buňka totiž nemá k dispozici vesmírnou stanici, v níž by si skafandr mohla svléknout. S buněčnou membránou se rodí, vyvíjí, množí i zaniká.
O existenci určité vrstvy na povrchu buněk, která je odděluje od vnějšího prostředí a má jiné vlastnosti než buněčný obsah, byli vědci přesvědčeni již od 19. století. Avšak první detailní pozorování umožnily až pokroky elektronové mikroskopie ve druhé polovině 20. století. Buněčná membrána (označovaná též jako cytoplazmatická či zkráceně plazmatická) je totiž natolik tenká, že pod optickým mikroskopem ji spatřit nelze. Samotná lipidová membrána (nepočítaje její případné další glykoproteinové „dekorace“) je silná zhruba 4 nm [1], což je asi 500× méně než velikost bakteriální buňky a 10 000× méně, než je velikost buňky eukaryotní. Objem, který tak plazmatická membrána drží pohromadě, je vskutku obdivuhodný a její význam je pro buňku zcela zásadní.
Jaké jsou další „pokročilé“ vlastnosti plazmatické membrány? Není nepropustná, umožňuje regulovanou výměnu látek – příjem zdrojů a vyloučení odpadu. Tím buňka udržuje stálé vnitřní prostředí odlišné od prostředí vnějšího, jinými slovy udržuje homeostázu. Plazmatická membrána je velmi proměnlivá v čase i prostoru a dokáže se přizpůsobit změně podmínek. Jejím prostřednictvím buňky komunikují, propojují se do tkání, pohybují se, poznávají se. Eukaryotické buňky využívají podobný typ membrány i na rozdělení svého vnitřního prostoru na organely (např. endoplazmatické retikulum, mitochondrie, lyzozomy, peroxizomy nebo buněčné jádro), čímž zvyšují efektivitu buněčných procesů a chemických reakcí potřebných k udržení homeostázy.
Základní stavební kameny: lipidy
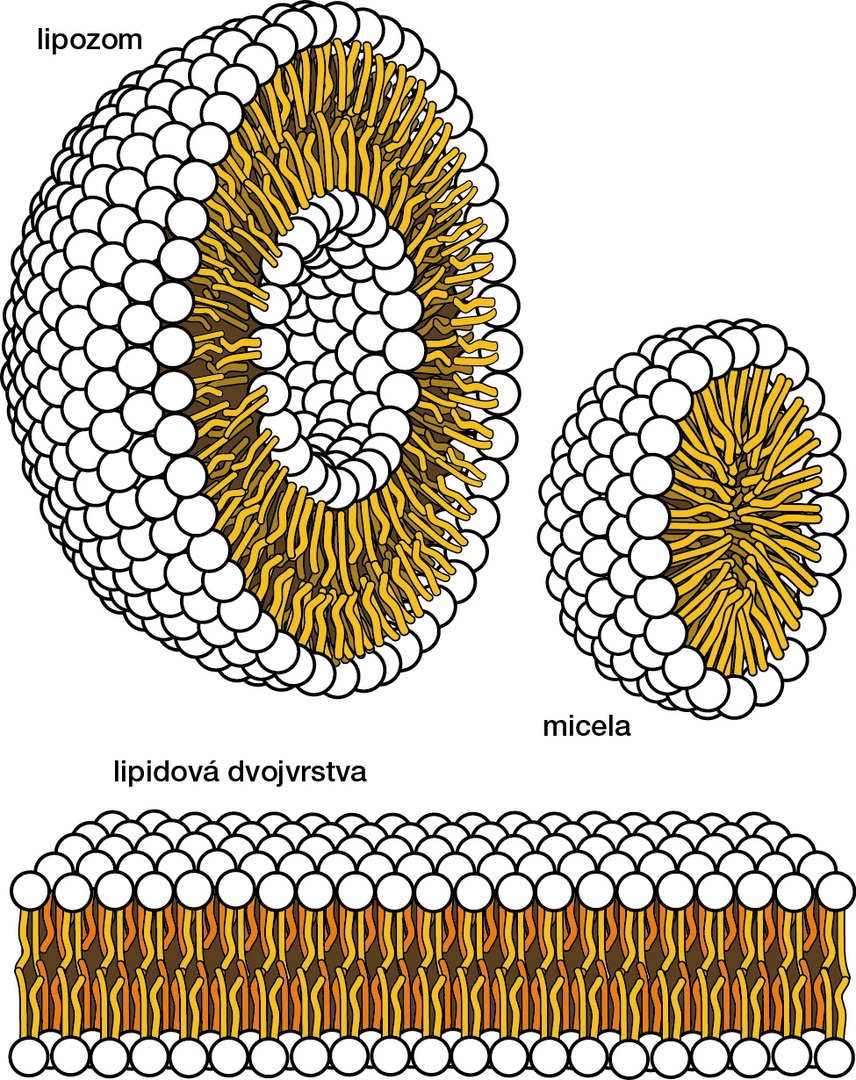
Abychom pochopili, jak buněčné membrány fungují, je třeba nejprve popsat, z čeho se skládají. Jejich hlavní složkou jsou lipidy, tedy tuky. Zásobní tuky, které známe třeba v podobě sádla či oleje, se skládají hlavně z triesterů mastných kyselin (karboxylových kyselin s dlouhým uhlovodíkovým řetězcem) a glycerolu, jsou to tedy triacylglyceroly (obr. 1A). Jsou tudíž nepolární a hydrofobní. Membránové lipidy jsou většinou estery diacylglycerolu a kyseliny fosforečné s vázanou další polární, hydrofilní skupinou (např. etanolamin, cholin, serin či glycerol), která se s vodou mísit může (obr. 1B, C) [2]. Ve výsledku jsou tedy membránové lipidy z jedné části hydrofobní a z druhé hydrofilní (obr. 1D), čili amfipatické. Proto když se ocitnou ve vodném prostředí (buňka je ze 70 % tvořena vodou), začnou se přirozeně orientovat a shlukovat tak, aby jejich hydrofilní část byla v kontaktu s vodným prostředím, zatímco hydrofobní část byla z vodného prostředí vyloučena. V závislosti na prostorovém tvaru lipidů a jejich množství pak vytvářejí útvary označované jako micely nebo lipidové dvojvrstvy, např. ve formě lipozomů (obr. 2).

Buňku si lze tedy zjednodušeně představit jako veliký „lipozom“, jehož membrána je tvořena dvěma vrstvami opačně orientovaných membránových lipidů (obr. 2). Pro lipidy je to uspořádání energeticky výhodné, za běžných podmínek se samy od sebe nerozpadnou, a fungují proto jako téměř nepropustná bariéra. Přes čistou lipidovou dvojvrstvu dokážou projít pouze plyny, malé hydrofobní molekuly (např. steroidní hormony) a v omezeném množství voda, tedy látky, které nenaruší strukturu membrány [2]. Vše, co je veliké či nabité, samovolně neprojde.
Fyzikální vlastnosti lipidových dvojvrstev jsou velmi závislé na teplotě [3]. Když se teplota zvyšuje, lipidy tají, jsou pohyblivější a lipidová membrána svým způsobem „teče“ (její složky vykazují laterální difuzi). Při hodně vysoké teplotě se dvojvrstva rozpadá. Naopak při snížení teploty membrány „tuhnou“ a lipidy v nich se stávají zcela nepohyblivými.
Pro svou optimální funkci buňka potřebuje udržet membránu tekutou, aby v ní probíhala laterální difuze, a tím mohlo docházet k chemickým reakcím potřebným pro homeostázu v membráně, k vratné asociaci lipidů mezi sebou a s proteiny (viz níže). U organismů neregulujících tělesnou teplotu to je docela oříšek; buňka musí být schopna na změnu teploty reagovat změnou nasycenosti a délky mastných kyselin, či obsahem lipidů výrazně měnících vlastnosti membrány (u živočišných buněk k nim patří např. cholesterol) [4].
U hypertermofilních organismů, např. některých druhů archeí, které žijí i při teplotách okolo 80–90 °C, je chemické složení buněčné membrány odlišné. Namísto mastných kyselin jsou k polární části lipidu připojeny izoprenoidní řetězce, které nejsou uprostřed membrány přerušeny (obr. 1E). Prakticky se tedy jedná o monovrstvu, která je mnohem stabilnější [5].
Funkční doplňky : proteiny
Druhou důležitou složkou buněčné membrány jsou proteiny s ní asociované. Ty tvoří v průměru asi 50 % její hmotnosti a dodávají membráně většinu jejích ostatních aktivních funkcí [4]. Vytvářejí kanály a přenašeče umožňující a regulující transport látek, které přes lipidy samy projít nemohou. Propojují buněčnou membránu s vnitřní kostrou buňky a s jejím okolím (extracelulární matrix), čímž zajišťují pevnost a soudržnost tkání. Zprostředkovávají komunikaci s okolím a signalizaci v buňce.
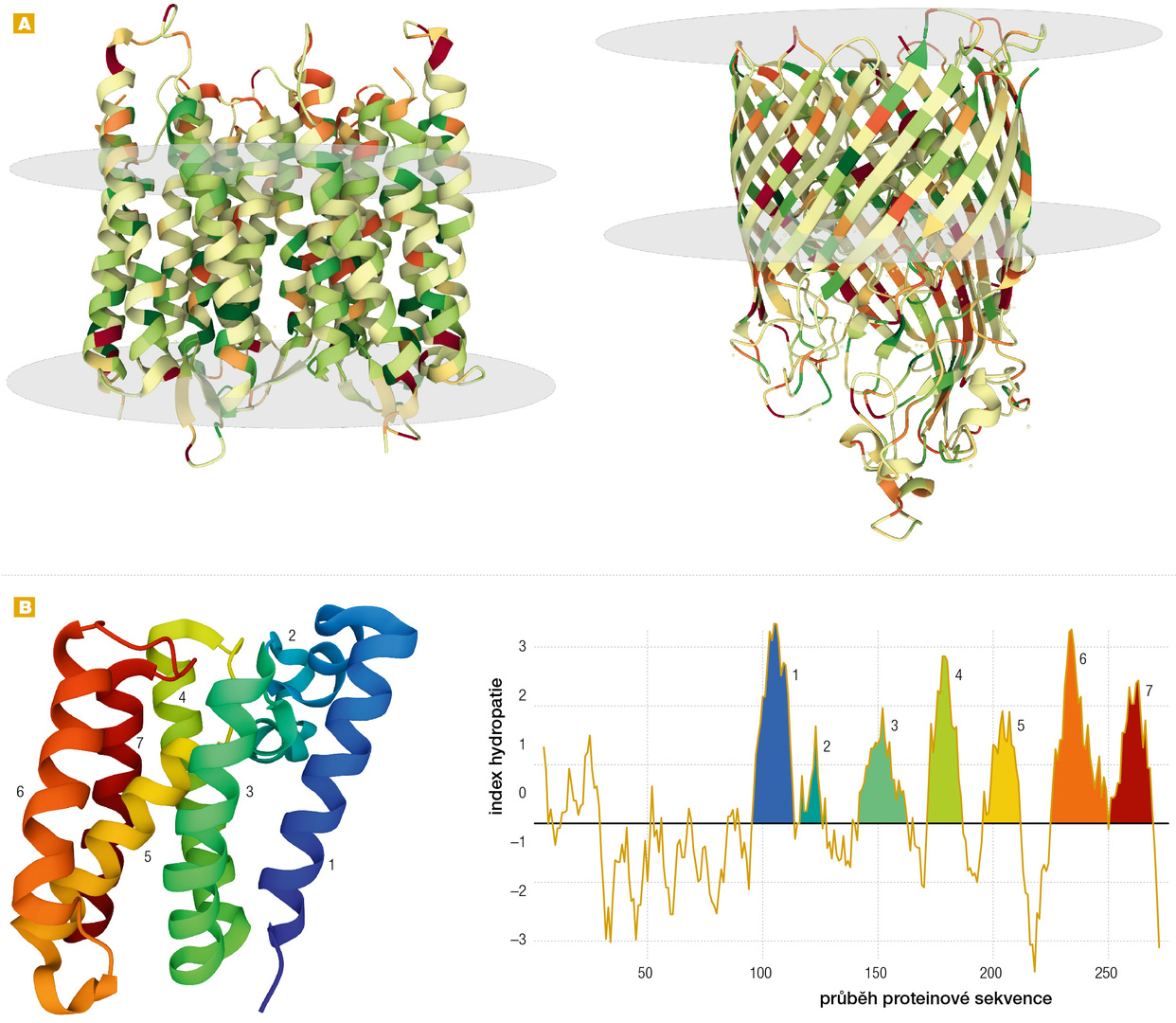
Odhaduje se, že zhruba čtvrtina všech buněčných proteinů je transmembránových. Svými vlastnostmi jsou odlišné od proteinů, které fungují ve vodném prostředí buňky [3, 4]. Obsahují souvislé úseky asi o 20–25 aminokyselinách s hydrofobními postranními řetězci (ve formě α-helixů či méně často β-řetězců), kterými procházejí skrz lipidovou dvojvrstvu. Na tyto úseky navazují hydrofilní části exponované do vodného prostředí buňky (obr. 3B). Při svém vzniku musí být transmembránové proteiny vkládány specializovanou proteinovou mašinérií, tzv. translokonem, rovnou do membrány (u eukaryot většinou endoplazmatického retikula, z něhož jsou poté i s membránou transportovány na místo určení) nebo dočasně udržovány v rozloženém stavu pomocí specializovaných proteinů, tzv. chaperonů, než budou do membrány vloženy. Jinak by se nesbalily do správné funkční struktury, agregovaly by a staly by se pro buňku toxickými.
„V jednom klasickém modelu je membrána popisována jako ‚fluidní mozaika‘:
lipidy představují moře a proteiny v nich plují jako ledové kry.“
Studium membránových proteinů je náročné. Jejich tvar a funkce jsou intimně spjaty s interfaciálním charakterem lipidové membrány a toto jejich přirozené prostředí není tak snadné simulovat. V jejich poznání jsme proto hodně pozadu v porovnání s ostatními buněčnými proteiny, které jsou volně rozpustné ve vodném prostředí.
Proměnlivost v prostoru a čase
Jak už bylo naznačeno, plazmatická membrána je dynamická a rozmanitá struktura. Její proměnlivost je dána již pouhou laterální difuzí lipidů a proteinů. V jednom klasickém modelu je membrána popisována jako „fluidní mozaika“: lipidy představují moře a proteiny v nich plují jako ledové kry. Proteiny i lipidy se mohou v membráně přechodně shlukovat a vytvářet různé funkční mikrodomény (Vesmír 91, 656, 2012/11), následně se rozpojovat a interagovat znovu někde jinde [3, 4].
Hlavním místem biosyntézy buněčných membrán je endoplazmatické retikulum (ER) [4]. V něm jsou syntetizovány tisíce různých lipidů i proteinů odlišných vlastností a funkcí, které obměňují membránu a přizpůsobují její složení aktuálním podmínkám. Plazmatická membrána si také udržuje asymetrii vrstev – lipidové složení vnější a vnitřní monovrstvy se liší. Díky fyzikálním vlastnostem membránových lipidů (polaritě, velikosti) se obě vrstvy samovolně nemíchají, respektive lipidy se napříč membránou pohybují s poločasem několika dnů, což z hlediska buňky nehraje velkou roli. Výjimkou je několik málo nepolárních lipidů (např. steroly či ceramid), které jsou schopny relativně rychlého transmembránového pohybu, tzv. flipování, v řádu sekund. Proteiny jsou rovněž příliš veliké a nabité, a nemohou se proto většinou v membráně „protočit“ zvenku dovnitř či naopak.
Vznik a udržení lipidové asymetrie katalyzují speciální enzymy, které za spotřeby energie „flipují“ lipidy. Roli hrají také interakce mezi lipidy a proteiny, důležité je i vnější pH. Porušení membránové asymetrie na plazmatické membráně vnímá zbytek organismu jako signál pro zničení buňky. Například expozice fostatidylserinu na vnější straně plasmatické membrány je znakem probíhající apoptózy a signálem pro fagocyty. Tento signál může umírající (např. infikovaná) buňka vydat prostřednictvím skramblázy – enzymu, který náhodně promíchá lipidy napříč dvojvrstvou [3].
Další variabilitu nacházíme při porovnání lipidového složení plazmatické membrány a membrán buněčných organel. Zdrojem těchto rozdílů je samotná biosyntéza strukturních lipidů v ER, Golgiho aparátu a na mitochondriích (a signálních lipidů i na jiných membránách) a jejich různě rychlý transport v membránovém systému pomocí váčků i přes kontaktní místa (viz níže). Například cholesterol je biosyntetizován v ER, ale jeho koncentrace tam jsou poměrně nízké, jelikož je efektivně transportován do plazmatické membrány, kde se akumuluje [6].
Významná je interakce membrán s buněčnou kostrou, tzv. cytoskeletem. Ten dokáže ve spolupráci s membránovými proteiny membránu vyklenout nebo vchlípit [2]. Tak vznikají buněčné výběžky, panožky, lamely, mikroklky. Plazmatická membrána buněk je ve skutečnosti plná takovýchto výběžků (obr. 4). Vyklenutou membránu je možné také odštěpit. Vzniknou malé samostatné váčky uvnitř či vně buňky (obr. 5).


Na principu váčků je založen buněčný membránový transport, protože při jejich odštěpení či splynutí se neporuší celistvý povrch buňky. Nepotřebné či poškozené části membrány jsou uklizeny do váčku a ten je posléze rozložen na základní stavební jednotky pomocí hydrolytických enzymů ve specializovaných membránových organelách, lyzozomech (Vesmír 95, 696, 2016/12). Podobným způsobem také buňky přijímají některé živiny, například železo či lipidy [2]. Pomocí membránových proteinů (receptorů) zachytí železo na molekule feritinu či lipoproteinovou částici a vzniklý komplex internalizují do váčku. V případě transferinového receptoru železa jsou schopné rozštěpit pouze náklad a receptor vrátit v neporušeném stavu na povrch, kde může dále plnit svoji funkci.
Váčkový transport plní signální funkci např. v nervové synapsi. Předpřipravené váčky splývají s membránou a do synaptického prostoru se rychle vylije velké množství neurotransmiteru [3].
Ve váčcích přicházejí na plazmatickou membránu a vnitrobuněčné membránové organely také nové membránové proteiny a lipidy syntetizované v endoplazmatickém retikulu. Nově se také ukazuje, že některé membránové organely (ER, plazmatická membrána, mitochondrie) jsou spolu spojeny v membránových kontaktních místech, ve kterých probíhá hlavně výměna lipidů mezi membránami za pomoci proteinů předávajících lipidy, které operují v mezimembránovém prostoru.
Významnou úlohu při obměně membránových proteinů hrají také proteolytické enzymy zakotvené na membráně nebo operující přímo uvnitř membrány (intramembránové) [3]. Mohou membránový protein rozstřihnout, a tím ho pomoci z membrány odstranit. Jejich působení může mít i signální funkci: část proteinu se ihned uvolňuje z buňky ven či dovnitř a spouští signální dráhu. Příkladem může být štěpení proteinu Notch, jehož prostřednictvím spolu buňky komunikují a rozrůzňují se během embryonálního vývoje živočichů. Notch receptor se při buněčné interakci propojí se svým protějškem na membráně druhé buňky, je „povytažen“ z membrány a dvě proteázy ho sekvenčně rozštěpí. Nejprve proteáza ADAM10 zakotvená na vnější straně plazmatické membrány odštěpí extracelulární část receptoru, který se tím dostává k druhé buňce a předává jí signál. Následně intramembránová aspartátová proteáza γ-sekretáza štěpí zbylou transmembránovou část receptoru uvnitř membrány a uvolňuje do nitra buňky proteinový fragment, který ovlivňuje genovou expresi buňky první. Ve výsledku tak obě buňky obdrží informaci, jak se mají dále vyvíjet.
Široké pole pro výzkum
Buněčná membrána je fascinující struktura, která je nezbytnou podmínkou existence všech živých organismů. Vznik polopropustné bariéry oddělující vnějšek od vnitřku byl v počátcích života stejně podstatný jako princip replikovatelné dědičné informace, dnes uložené v molekulách nukleových kyselin [7]. Biologické membrány určitých vlastností umožňují buňkám přežívat i ve velmi nehostinném prostředí, komunikovat a být součástí mnohobuněčných organismů. Výzkum na tomto magickém rozhraní se dnes zaměřuje zejména na pochopení mechanismu regulace homeostázy v biologické membráně, na principy biogeneze a obměny membránových proteinů a jejich funkce. Přináší poznatky využitelné pro pochopení počátků života i léčbu řady chorob, od infekčních po moderní civilizační včetně metabolických a neurodegenerativních. Ale o tom více někdy příště.
Autoři děkují Dr. Marku Vrbackému za výběr úvodního citátu.
Literatura
[1] Mitra K. et al.: Modulation of the bilayer thickness of exocytic pathway membranes by membrane proteins rather than cholesterol. Proc. Natl. Acad. Sci. U. S. A. 101, 4083–4088, 2004, DOI: 10.1073/pnas.0307332101.
[2] Stillwell W.: An Introduction to Biological Membranes. Elsevier, 2013.
[3] Luckey M.: Membrane structural biology. Cambridge University Press, 2008.
[4] Yeagle P. L.: The Structure of Biological Membranes. CRC Press, 2012.
[5] van de Vossenberg J. L. et al.: The essence of being extremophilic: the role of the unique archaeal membrane lipids. Extremophiles 2, 163–170, 1998, DOI: 10.1007/s007920050056.
[6] van Meer G. et al: Membrane lipids: where they are and how they behave. Nat. Rev. Mol. Cell Biol. 9, 112–124, 2008, DOI: 10.1038/nrm2330.
[7] Kahana A., Lancet, D.: Self-reproducing catalytic micelles as nanoscopic protocell precursors. Nat. Rev. Chem. 5, 870–878, 2021, DOI: 10.1038/s41570-021-00329-7.
Ke stažení
 článek ve formátu pdf [1,34 MB]
článek ve formátu pdf [1,34 MB]
O autorech
Šárka Žaloudková Boháčová
Kvido Stříšovský
Další články k tématu
Iluze prostoru

Do hloubky povrchnosti
(Ne)smáčivost inspirovaná přírodou
Zemský povrch na mapách
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů
