









Čtvrtá vrstva
 Zárodečné vrstvy – tedy buněčné embryonální vrstvy, ze kterých během individuálního vývoje vznikají tkáně a orgány – jsou hned vedle buněk považovány za základ strukturální organizace embrya každého mnohobuněčného živočicha. První pozorování zárodečných vrstev či listů pochází již z počátku 19. století (viz rámeček 2) a schémata vzniku a vývoje těchto vrstev jsou dodnes tím nejzákladnějším, co si odnášíme z hodin obecné biologie. Standardně rozeznáváme tři zárodečné vrstvy – ektoderm, entoderm a mezoderm – a právě podle jejich přítomnosti se mnohobuněční živočichové (Metazoa) učebnicově rozdělují na Monoblastika, Diblastika a Triblastika.
Zárodečné vrstvy – tedy buněčné embryonální vrstvy, ze kterých během individuálního vývoje vznikají tkáně a orgány – jsou hned vedle buněk považovány za základ strukturální organizace embrya každého mnohobuněčného živočicha. První pozorování zárodečných vrstev či listů pochází již z počátku 19. století (viz rámeček 2) a schémata vzniku a vývoje těchto vrstev jsou dodnes tím nejzákladnějším, co si odnášíme z hodin obecné biologie. Standardně rozeznáváme tři zárodečné vrstvy – ektoderm, entoderm a mezoderm – a právě podle jejich přítomnosti se mnohobuněční živočichové (Metazoa) učebnicově rozdělují na Monoblastika, Diblastika a Triblastika.
V tomto článku však budeme uvažovat názor odlišný. My obratlovci se od ostatních skupin odlišujeme hlavně novými tkáněmi, které jsou většinou odvozené od buněk neurální lišty. Obratlovci vznikli díky zásadním přeměnám embryogeneze. Buňky neurální lišty přebudovávají zavedená schémata triblastického vývoje, v němž nově hrají zásadní roli indukční procesy a epigenetické interakce. Buňky neurální lišty mohou být lehce považovány za další zárodečnou vrstvu a obratlovci za živočichy nikoliv triblastické, ale tetrablastické.1)
Zárodečné vrstvy a „stavební plán“ obratlovců
 Zárodečné vrstvy představují jeden ze základů pochopení organizace živočišného těla. U „vyšších živočišných skupin“ vznikají povrchové struktury související s pokožkou a nervovým systémem z vnější zárodečné vrstvy, ektodermu. Z vnitřní vrstvy, entodermu, vznikají soustavy orgánů uvnitř našeho těla, jako je např. trávicí soustava či trávicí žlázy. Všechny zbylé tkáně (kosti, svaly, cévní či močopohlavní soustava) vznikají z mezodermu. Toto klasické učebnicové rozdělení však překvapivě neplatí pro obratlovce, přestože jsou mezi triblastické kmeny počítáni. Kupříkladu v hlavě obratlovců je drtivá většina skeletotvorných a pojivových tkání embryonálně odvozena z buněk neurální lišty. Buňky neurální lišty tvoří i významnou část soustavy nervové a přispívají do mnoha vnitřních orgánů.2)
Zárodečné vrstvy představují jeden ze základů pochopení organizace živočišného těla. U „vyšších živočišných skupin“ vznikají povrchové struktury související s pokožkou a nervovým systémem z vnější zárodečné vrstvy, ektodermu. Z vnitřní vrstvy, entodermu, vznikají soustavy orgánů uvnitř našeho těla, jako je např. trávicí soustava či trávicí žlázy. Všechny zbylé tkáně (kosti, svaly, cévní či močopohlavní soustava) vznikají z mezodermu. Toto klasické učebnicové rozdělení však překvapivě neplatí pro obratlovce, přestože jsou mezi triblastické kmeny počítáni. Kupříkladu v hlavě obratlovců je drtivá většina skeletotvorných a pojivových tkání embryonálně odvozena z buněk neurální lišty. Buňky neurální lišty tvoří i významnou část soustavy nervové a přispívají do mnoha vnitřních orgánů.2)
 Neurální lišta (obr. 1 a obr. 2) je často popisovaná jako přechodná embryonální struktura, vznikající během neurulace na hřbetní straně embrya. Během vzniku neurální trubice se z tohoto materiálu postupně oddělí obrovské množství buněk, které migrují k břišní straně a vytvářejí spoustu nových buněčných derivátů. Zároveň přispívají do struktur, které u původnějších skupin vznikají z odlišných zárodečných vrstev (viz rámeček 1). Kromě buněk neurální lišty se obratlovci liší od ostatních skupin i populací epidermálních plakod, které vytvářejí základy všech smyslových orgánů i část nervové soustavy a společně s buňkami neurální lišty dávají vznik orgánům odpovědným za interakce s okolím. Obratlovci tedy nemohou být považováni za klasická triblastika. Od ostatních skupin se evolučně oddělili právě tím, že si z buněk neurální lišty vytvořili mnoho nových buněčných typů, tkání a interakcí, jež tvoří evoluční nadstavbu nad původním stavebním plánem triblastik. Můžeme však tyto tkáně považovat za další zárodečný list?
Neurální lišta (obr. 1 a obr. 2) je často popisovaná jako přechodná embryonální struktura, vznikající během neurulace na hřbetní straně embrya. Během vzniku neurální trubice se z tohoto materiálu postupně oddělí obrovské množství buněk, které migrují k břišní straně a vytvářejí spoustu nových buněčných derivátů. Zároveň přispívají do struktur, které u původnějších skupin vznikají z odlišných zárodečných vrstev (viz rámeček 1). Kromě buněk neurální lišty se obratlovci liší od ostatních skupin i populací epidermálních plakod, které vytvářejí základy všech smyslových orgánů i část nervové soustavy a společně s buňkami neurální lišty dávají vznik orgánům odpovědným za interakce s okolím. Obratlovci tedy nemohou být považováni za klasická triblastika. Od ostatních skupin se evolučně oddělili právě tím, že si z buněk neurální lišty vytvořili mnoho nových buněčných typů, tkání a interakcí, jež tvoří evoluční nadstavbu nad původním stavebním plánem triblastik. Můžeme však tyto tkáně považovat za další zárodečný list?
Primární a sekundární zárodečné vrstvy

 K formování finálních zárodečných vrstev dochází až během gastrulace (viz rámeček 3). Ektoderm a entoderm jsou však jako předběžné vrstvy přítomny již ve vajíčku před oplozením, kde se formují díky informaci zděděné po matce. Pro jejich vznik není nutná interakce pohlavních buněk. Ektoderm a entoderm jsou primární zárodečné vrstvy, neboť se v ontogenezi zakládají nejdříve a jsou fylogeneticky původnější – monoblastika a diblastika předcházela triblastikům. Mezoderm se objevuje až po oplodnění a jeho embryonální vznik z ektodermu nebo z pomezí ekto-entodermu je indukován tkáňovými interakcemi. Mezoderm je zárodečná vrstva odvozená ontogeneticky i evolučně – je to sekundární zárodečná vrstva. Neurální lišta může být považována za další sekundární zárodečný list ze stejných důvodů. Obě vznikaji velmi časně a obě díky induktivním interakcím mezi primárními listy (buňky neurální lišty vznikají na pomezí neurálního a nonneurálního ektodermu kaskádou reakcí zvanou neurální indukce).
K formování finálních zárodečných vrstev dochází až během gastrulace (viz rámeček 3). Ektoderm a entoderm jsou však jako předběžné vrstvy přítomny již ve vajíčku před oplozením, kde se formují díky informaci zděděné po matce. Pro jejich vznik není nutná interakce pohlavních buněk. Ektoderm a entoderm jsou primární zárodečné vrstvy, neboť se v ontogenezi zakládají nejdříve a jsou fylogeneticky původnější – monoblastika a diblastika předcházela triblastikům. Mezoderm se objevuje až po oplodnění a jeho embryonální vznik z ektodermu nebo z pomezí ekto-entodermu je indukován tkáňovými interakcemi. Mezoderm je zárodečná vrstva odvozená ontogeneticky i evolučně – je to sekundární zárodečná vrstva. Neurální lišta může být považována za další sekundární zárodečný list ze stejných důvodů. Obě vznikaji velmi časně a obě díky induktivním interakcím mezi primárními listy (buňky neurální lišty vznikají na pomezí neurálního a nonneurálního ektodermu kaskádou reakcí zvanou neurální indukce).
Jak mezoderm, tak neurální lišta dávají vznik embryonálnímu mezenchymu (embryonální podpůrné tkáni). To ukazuje, že při vzniku obratlovců převzala neurální lišta značnou část funkcí mezodermu. Z evolučního hlediska znamenal vznik mezodermu obrovskou změnu v časoprostorovém přístupu k původním tělním plánům diblastik a vedl k rozrůznění přibližně dvaceti nových živočišných skupin, které souhrnně nazýváme triblastické (od žahavců po strunatce). Vznik další sekundární zárodečné vrstvy, neurální lišty, vedl k diferenciaci obratlovců a k dalším nedozírným změnám zavedených vývojových mechanismů.
Neurální lišta jako základní embryonální specifikum obratlovců
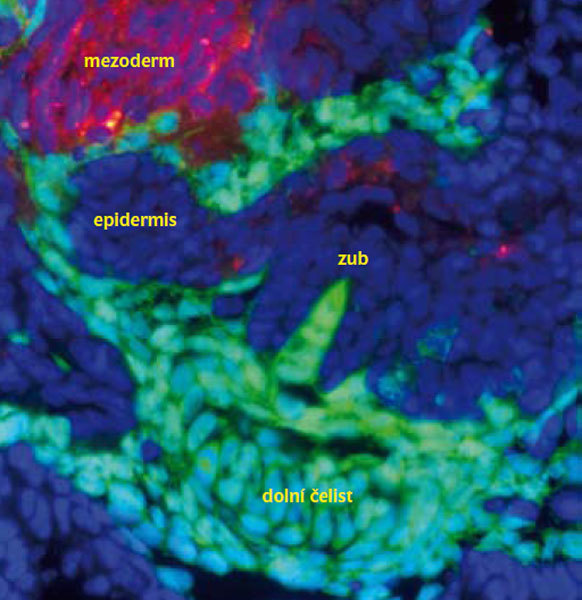
Většina znaků, jimiž se obratlovci odlišují od ostatních skupin (hlavové nervy se senzorickými ganglii, přední mozek, párové modifikace hltanu (faryngu) a jeho skeletálních elementů, pokryvný skeletální systém ap.) je z větší části odvozena od buněk neurální lišty. Z embryonálního hlediska je tedy neurální lišta nejdůležitějším původním znakem obratlovců. Právě ona embryonálně konfiguruje základní evoluční novotvar obratlovců – hlavu.3) Zatímco v trupu migrují buňky neurální lišty jednotlivě, v hlavě vytvářejí tři souvislé buněčné proudy4) s obrovským množstvím buněk (obr. 2). Typickou vlastností takto nově vzniklých buněk je semikmenovost čili multipotence. To znamená schopnost adaptivně reagovat2) a rozlišit se do obrovského množství buněčných typů a derivátů. Plasticitu a rozdíly ve skeletogenní potenci mezodermu a buněk neurální lišty zkoumal následující experiment, při němž byla část hlavového mezodermu nahrazena buňkami neurální lišty.5) Vznikla experimentální kuřata, u nichž část mezodermální lebky bez jakýchkoliv pozorovatelných problémů vznikla z buněk neurální lišty, které se ukázaly být vývojově schopné mezoderm plně nahradit. Evoluční úspěch obratlovců je z části přisuzován právě této vývojové plastičnosti našich hlavových tkání. Evoluční vznik zubů a čelistí6) či ptačího zobáku7) – to vše a mnohé další vzniklo díky plastičnosti mezenchymu odvozeného od buněk neurální lišty a jeho schopnosti adaptivně reagovat.
Zárodečné vrstvy, homologie a evolučně-vývojová biologie
Od objevení zárodečných vrstev r. 1817 se považovalo za samozřejmé, že homologickéorgány musejí bezpodmínečně vznikat ze stejných zárodečných vrstev. Anatomické homologie, hledané do té doby pouze na dospělcích, se transformovaly do homologií embryonálních. Embrya se stala důkazem evoluce a embryonální procesy začaly být nahlíženy jako příčiny evolučních změn (od původního náhledu „ontogeneze rekapituluje fylogenezi“ k modernějšímu „sled ontogenezí vytváří fylogenezi“). Ukázalo se také, že struktury klasicky považované za homologické napříč skupinami nemusejí nutně embryonálně pocházet ze stejné zárodečné vrstvy a nemůžeme ani očekávat, že homologické elementy vyrůstají cestou identických vývojových mechanismů. Homologie, čili kontinuita vývojové informace od předka k následovníkům, je mnohoúrovňový a značně nelineární problém. K evolučním změnám může dojít na kterékoliv úrovni hierarchicky uspořádaných vývojových programů a hledání „neviditelných linek plození, které nás spojují úplně se vším živým (jak by řekl Darwin)“,8) je nesmírně obtížné. Posuďte sami: kosti a chrupavky v přední části hlavy obratlovců vznikají z buněk neurální lišty, zatímco zadní část stejných hlavových tkání vzniká z mezodermu. Obdobně si většina druhoústých živočichů s hltanovými štěrbinami vytvořila podpůrný skelet těchto štěrbin, který je však u původních skupin tvořen pouze nebuněčnou chrupavkou a je vylučován výhradně epiteliálními buňkami ektodermu, entodermu a mezodermu. U obratlovců ale nacházíme v hltanu struktury, které jsou tvořeny kompaktními buněčnými elementy, jež mají původ v migrujících buňkách neurální lišty.9) Kontinuita informace (zde např. exprese SoxE genů řídících morfogenezi kolagenů, proteinů typických pro chrupavku) nám však jasně naznačuje, že tu jde o vzájemně homologické struktury, přestože jejich embryonální původ a buněčná struktura tkání během evoluce prošly zásadními změnami.
Ch. Darwin a A. R. Wallace ukázali, že na evoluci můžeme pohlížet jako na „společný původ s následnými modifikacemi“. Moderní syntéza 30. a 40. let minulého století viděla evoluční změny převážně jako statistickou populační dynamiku, zatímco po objevu struktury DNA byla evoluce vnímána prizmatem dědičných změn v sekvencích DNA. Evolučně-vývojová biologie10) se zaměřuje na zkoumání otázek, jak se ontogeneze vyvíjí v průběhu evoluce nebo které změny ve vývojových procesech ovlivňují (či vytvářejí) evoluci. Pohled vědy evo-devotické11) nám také umožňuje nahlížet obratlovce jako organismy, které evolučně vznikly díky zásadnímu přebudování triblastického vývoje (hlavně) pomocí buněk neurální lišty, které mohou být následně považovány za další zárodečnou vrstvu a obratlovci za organismy tetrablastické.
Poznámky
1) Např. B. K. Hall: Evolution and Development 2(1), 3–5, 2000; B. K. Hall: The Neural Crest and Neural Crest Cells in Vertebrate Development and Evolution. Springer, 2009
2) N. M. LeDouarin a C. Kalcheim: The Neural Crest. Cambridge University Press, 1999
3) Např. R. G. Northcutt: Journal of Experimental Zoology (Mol. Dev. Evol.) 304B, 274–297, 2005
4) Např. R. Cerny et al.: Developmental Biology 266, 252–269, 2004
5) R. A. Schneider: Developmental Biology 208, 441–455, 1999
6) V. Soukup, H. Epperlein, I. Horáček & R. Cerny: Nature 455(7214), 795–798, 2008; R. Cerny et al.: A new model for the evolution of the vertebrate jaw (submitted 2010)
7) Např. R. A. Schneider & J. A. Helms: Science 299, 565–568, 2003; A. Abzhanov: Science 305, 1462–1464, 2004
8) Viz úvodník Davida Storcha,
Vesmír 88, 431, 2009/7
9) A. L. Rychel a B. J. Swalla: Journal of Experimental Zoology (Mol. Dev. Evol.) 308B, 1–11, 2007
10) B. K. Hall: Evolutionary Developmental Biology. Kluwer, 1999
11) viz také A. Markoš, Vesmír 88, 593, 2009/9
Co vzniká z buněk neurální lišty
V hlavě obratlovců vytvářejí buňky neurální lišty drtivou většinu skeletálních tkání: chrupavky, kosti, buňky pojivových tkání či škáru, mimo sklovinu všechny tkáně zubu. Také všechny pojivové tkáně hlavových svalů či tkáně, kam se tyto svaly upínají, mají původ v neurální liště. Vznikají z ní všechny pigmentové buňky celého těla (melanocyty, xantophory, erythrophory či iridiophory), nervové buňky (glie, parasympatické i sympatické neurony, enterické či senzorické neurony, Schwannovy buňky atd.), část předního mozku a větší část periferního nervstva (zbylá část vzniká z plakod). Přispívají i do kardiovaskulárního systému, cév, buněk endokrinního systému atd. Přestaneme-li se tedy na buňky neurální lišty dívat jako na několik podivně migrujících buněk a pokusíme-li se o vhled odpovídající současným poznatkům, uvidíme mohutnou a významnou populaci (obr. 2), dávající vznik většině tkání hlavy obratlovců.
Historie objevů
Christian H. Pander: Roku 1817 objevil zárodečné vrstvy na kuřecím embryu, nazval je Serosenblatt, Schleinblatt a Gefässblatt.
Martin Rathke: Objevil na korýši r. Astacus zárodečné vrstvy podobné kuřecím (1825, 1829).
K. E. Von Baer: „Otec srovnávací embryologie“. Rozšířil Panderovo poznání o zárodečných vrstvách na všechny obratlovce (1828). Embryo se vyvíjí od obecného k speciálnímu.
George J. Allman: Profesor přírodních věd v Edinburgu (1855–1870). Roku 1853 poprvé použil dnešní termíny ektoderm a entoderm. Pro něj to však byly pouze odlišné názvy částí dospělého žahavce.
Robert Remak (1815–1865): První rozpoznal mezoderm jako další vrstvu zárodku kuřete (1950–1955). Odhalil také odlišnou histologickou charakteristiku každé vrstvy.
Thomas Huxley: Jako první zhomologizoval ekto- a entoderm bezobratlých a obratlovců (1849). Pro střední vrstvu razil termín mezoderm (1871). Rozšířil poznání o zárodečných vrstvách z embryologie a homologie do fylogeneze. Nutno však mít stále na paměti, že T. Huxley, stejně jako jeho kolegové, neuvažoval v dnešním slova smyslu evolučně. Vyjma Darwina a pár dalších výjimek nikdo v té době nemyslel na vznik „odvozenějších“ skupin z „nižších“. Stále silně převažovalo Cuvierovo mínění: organismy jsou odlišné, oddělené skupiny bez vzájemných fylogenetických vazeb (Vesmír 89, 164, 2010/3). Přesto, nebo právě proto, je Huxleyho mínění o homologizaci ektodermu a entodermu obratlovců s vnější a vnitřní vrstvou těla bezobratlých tak důležité.
Wilhelm His (1831–1904): Švýcarský embryolog, profesor anatomie a fyziologie v Basileji. První popsal neurální lištu jako vrstvu mezi neurálním a epidermálním ektodermem (1868) na zárodku kuřete. Nazval ji Zwischenstrang. Pro zajímavost: Objevil místo, kde byl pohřben J. S. Bach a identifikoval jeho lebku srovnáním lebky nalezené s jílovým modelem, vyrobeným dle oficiálního portrétu. Význačný oponent Hackelova biogenetického zákona, podle kterého ontogeneze rekapituluje fylogenezi.
Ernst Haeckel (Vesmír 84, 712, 2005/12): Snažil se nalézt odpověď, proč se raná embrya recentních obratlovců podobají dospělci kopinatce (Branchiostoma), viz rovněž např. Vesmír 76, 89, 1997/2 a 78, 509, 1999/9. Odpověď dala jeho teorie Gastraea (1872, 1874), spojující poznatky z morfologie a homologie, embryologie a evoluce. Podle ní se všichni mnohobuněční živočichové vyvinuli z jednoho předka, jak na to ukazuje stadium blastuly. Tohoto předka pojmenoval právě Gastraea. Stopování předků (často dle znalostí embryologie) umožnilo Hackelovi konstruovat fylogenetické stromy příbuznosti.
Francis M. Balfour (1851–1882): Jeden z nejlepších embryologů vůbec. Ačkoliv zemřel velmi mladý při výstupu v Alpách, zanechal po sobě obrovské množství cenných pozorování. Již ve 22 letech rozpoznal, že ne vše v kuřecím embryu nám může poskytnout fylogenetickou informaci a jako první rozeznal sekundární embryonální adaptace a mluvil o Darwinistické selekci již v raných fázích vývoje embrya.
E. R. Lancaster: Roku 1877 rozdělil živočichy (dle znalostí zárodečných vrstev, které označoval Balfourovými termíny ekto-, meso- a hypoblast) na Homoblastica, Diploblastica a Triploblastica, což se udrželo více méně do dnešních dnů. Rozšířil tedy teorii zárodečných vrstev i do systematiky.
Oskar a Richard Hertwigové: Definovali mezenchym (1882) jako buňky opouštějící mezoderm a tvořící pojivové tkáně a krev.
Adam Sedgwick: Balfourův nástupce v Cambridge, asi nejvýznačnější kritik teorie zárodečných vrstev (1894, 1895). Jeho kritika, minimálně z části oprávněná, vyvstávala z jeho vlastních přesných pozorování původu mezodermu, periferního nervstva a neurální lišty.
Julia Platt: Ve svých pracech (1893, 1894) prokázala, že chrupavky hltanu obratlovců, stejně jako kraniofaciální skelet, mají původ v mezenchymu neurální lišty a (jak věřila) i plakod. Ačkoliv její pionýrské práce byly velmi přesvědčivé a podporovali je i někteří další kolegové, byly přes 50 let přehlíženy. Studovala na Harvardu, v Chicagu a Mnichově a byla jednou z prvních žen, kterým se podařilo získat doktorát na německé univerzitě ve Freiburgu (1898). I přes dvanáct jejích významných prací se jí však nepodařilo nalézt místo na jakékoliv univerzitě, zčásti možná kvůli otevřené kritice jejích mužských kolegů.
F. L. Landacre a L. S. Stone: Přinesli další důkazy pro původ mnoha tělních a hlavových struktur z neurální lišty (1921– 1930) a opravili některé mylné interpretace Plattové, týkající se materiálu ektodermálních plakod.
sir Gavin R. deBeer: Profesor embryologie v Londýně a vedoucí Britského Muzea. Zpochybnil teorii zárodečných vrstev (1947) s poukazem na to, že orgány, homologické u různých živočichů, nepovstávají ze stejných vrstev. Svými pracemi význačně přispěl k poznání derivátů neurální lišty.
Sven O. Hörstadius (1898–1996): Profesor zoologie v švédské Uppsale, význačný embryolog a ornitolog, čestný člen mnoha nejstarších univerzit světa. Autor první (1950) a velmi významné knižní monografie o neurální liště (zvané dnes často jednoduše „Hörstadius“).
Brian K. Hall: Profesor na Dalhousie University v Halifaxu v Kanadě. Jeden z dnešních nejvýznačnějších zastánců neurální lišty jako čtvrté zárodečné vrstvy. Autor knih Evolutionary Developmental Biology (1992, 1998), Neural Crest in Development and Evolution (1999, 2009); editor knihy Homology: The Hierarchical Basis of Comparative Biology (1994).
Formování zárodečných vrstev
Během gastrulace se díky morfogenetickým pohybům a změnám v expresi genů přeskupují buňky a utvářejí finální vrstvy zárodečných buněk, tzv. zárodečných listů, které známe jako ektoderm, entoderm a mezoderm. Důkazem je i fenomén zvaný selektivní buněčná afinita. Jestliže před gastrulací šetrně rozmělníme předchozí vývojové stadium (blastulu) na jednotlivé buňky a držíme je v kultivačním médiu, budou se náhodně shlukovat s jim nejbližšími buňkami. Provedeme-li však stejný pokus se zárodkem ve stadiu pozdní gastruly, buňky se začnou shlukovat selektivně podle příslušnosti ke konkrétní zárodečné vrstvě, ze které pocházejí. Ektodermální, entodermální a mezodermální buňky utvoří vždy jasně odlišné vrstvy, čímž nám vznikne kompletní gastrula de novo. Zatímco tedy v blastule jsou přítomny jen zárodečné vrstvy předběžné, v pozdní gastrule již mají charakter pravých zárodečných listů.
Ke stažení
 článek ve formátu pdf [787,02 kB]
článek ve formátu pdf [787,02 kB]
O autorovi
Robert Černý
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
