









Táto, mámo, máme molu!
| 20. 4. 2018O početí nového lidského života často mluvíme jako o fascinujícím zázraku. Mechanismy tvorby pohlavních buněk a oplození jsou však evolucí vyšroubovány k takové složitosti, že se celý proces ne vždy povede. Někdy chyba vede k neplodnosti, někdy k vážným vrozeným poruchám typu Downova syndromu – a někdy ke vzniku zvláštních, hrubě aberantních produktů početí, které se vymykají klasickým zákonitostem lidské dědičnosti. Do života rodičů přinášejí tyto genetické entity smutek, z hlediska forenzní DNA analýzy jsou však nesmírně zajímavé a lze na nich ukázat smysl a půvab použití bayesovské inference při hodnocení důkazu.
Jeden každý z laskavých čtenářů a jedna každá z laskavých čtenářek těchto řádků započali svou existenci spojením dvou pohlavních buněk: otcovy haploidní (= jednu sadu chromozómů nesoucí) spermie a matčina haploidního vajíčka. Obě chromozomální sady po oplození vytvořily společné jádro nově vzniklé zygoty, tedy první diploidní (= dvě sady chromozómů nesoucí) buňky, jejímž dělením a diferenciací pak čtenář či čtenářka vznikli. Snad nebudou vyděšeni či dotčeni, prozradíme-li jim, že z hlediska genetické klasifikace a nomenklatury jsou takzvaným biparentálním monogynickým monoandrickým diploidem1) neboli normoploidem.
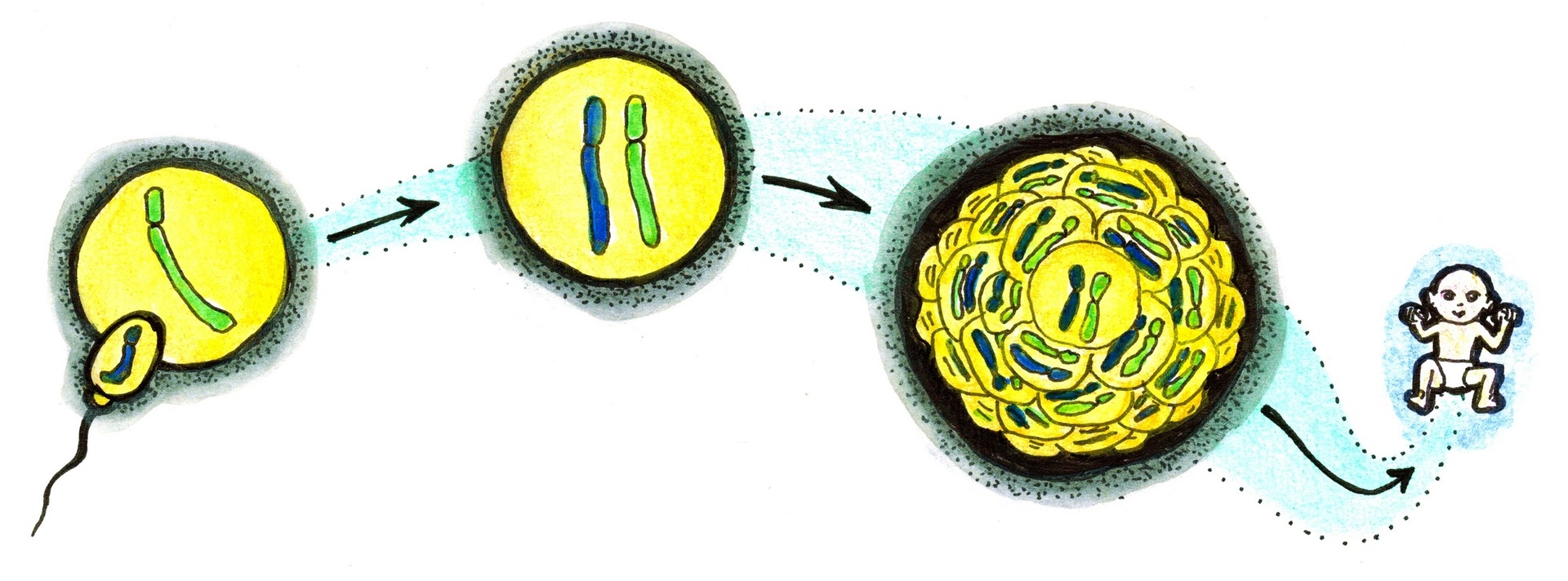
Normální vývoj lidského embrya zajišťuje a naději na vznik zdravého jedince dává právě pouze přísná diploidie, při níž jsou všechny autozómy, tj. nepohlavní chromozómy č. 1–22, přítomny v buněčném jádru po dvou – mluvíme o jejich disomii. Už odchylka v počtu jednotlivého autozómu vede v lepším případě k závažnému patologickému stavu (například rozvoji již zmíněného Downova syndromu při trisomii 21. chromozómu), v tom horším (a častějším) ke spontánnímu potratu nebo k velmi brzkému úmrtí novorozence.
Na vině je zejména neschopnost buněk efektivně kompenzovat genovou dávku daného autozómu, tedy při nadbytečném chromozómu úměrně snížit a při chybějícím chromozómu naopak úměrně zvýšit expresi stovek na něm ležících genů tak, aby celková dávka odpovídala normě. Kompenzace je buňka relativně dobře schopna pouze při odchylkách v počtu gonozómů, tj. pohlavních chromozómů X a Y.2)
Zdálo by se tedy, že změna počtu celých chromozomálních sad musí zákonitě vést k chaosu v buňce a k její neschopnosti dlouhodobější existence, ovšem tak jednoduché to není. Pojďme se podívat, co všechno se může pokazit a jaké důsledky to má.
Spermie v prázdném domě
Chybou během tvorby vajíček neboli oogeneze může vzniknout prázdné, bezjaderné vajíčko, v němž není přítomen žádný genetický materiál – je nulliploidní. Přesto ale může být oplodněno normální haploidní spermií, která nese 23 chromozómů (22 autozómů a jeden gonozóm, X nebo Y). Haploidie jako taková sice není slučitelná s dalším vývojem, existuje však obezlička: v jádře vajíčka může dojít k duplikaci chromozómů bez toho, že by se současně celé vajíčko rozdělilo na dvě dceřiné buňky – proběhne tzv. endoreduplikace). Tím se vajíčko stane diploidním – mluvíme o uniparentální nulligynické diandrické diploidii.
Pokud spermie nesla pohlavní chromozóm Y, je konec: jakkoli si totiž vajíčko endoreduplikací pomohlo k diploidii, má sestavu pohlavních chromozómů YY a bez chromozómu X je jeho další vývoj nemožný. Pokud ale spermie nesla chromozóm X, má vajíčko sestavu pohlavních chromozómů XX, a tedy zdánlivě vše potřebné k dalšímu vývoji.
Diploidní buňka může z prázdného vajíčka ovšem vzniknout i jinak, a to tzv. dispermií, tedy oplozením nikoli jednou, ale dvěma spermiemi. Rovněž tady platí, že tyto spermie musejí nést pohlavní chromozómy X a Y nebo X a X, aby oplozené vajíčko mohlo pokračovat ve vývoji.
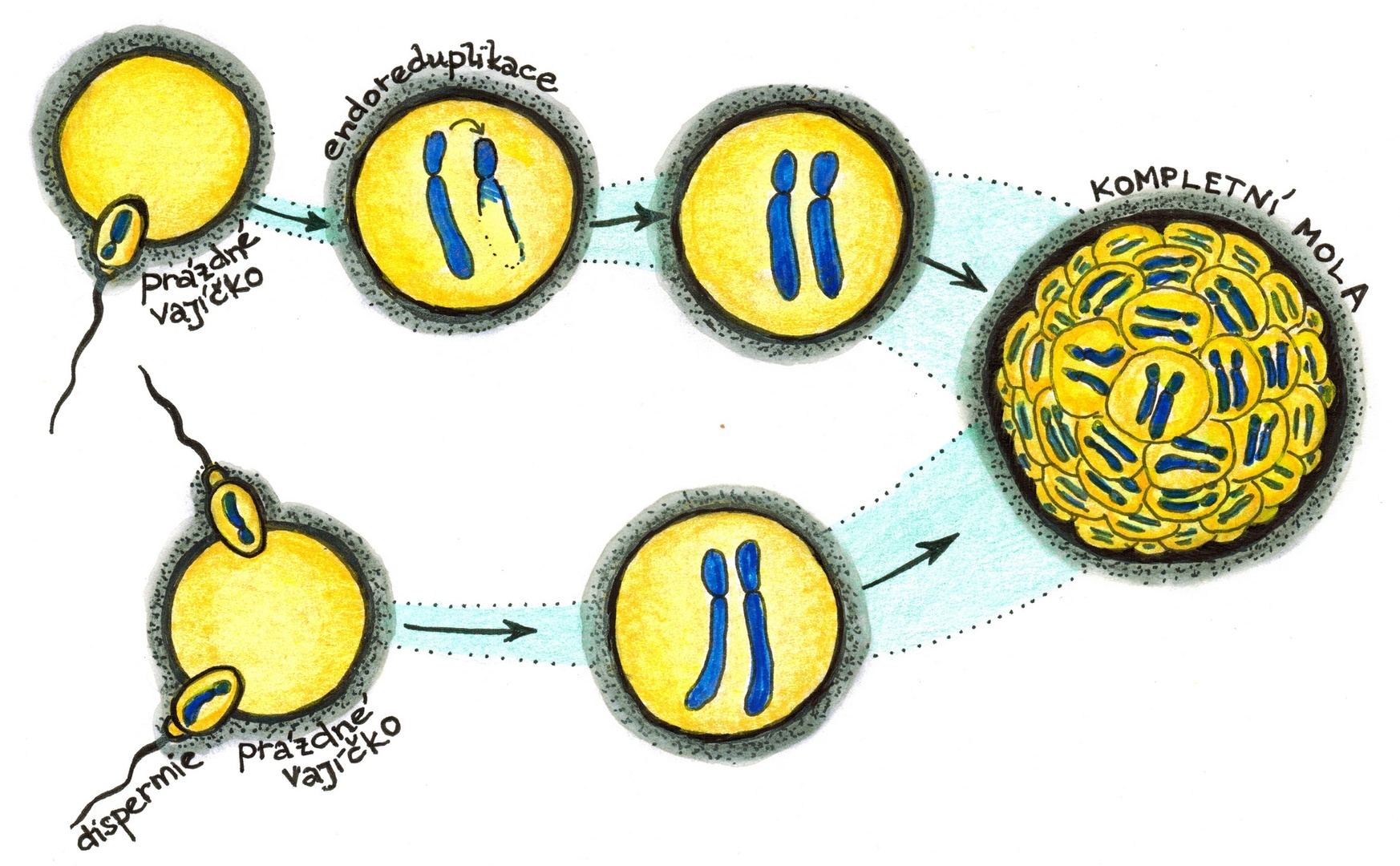
Diandrický diploid se dělí zpravidla rychle a zběsile, přičemž dává vzniknout mohutně rozvinutému patologicky organizovanému trofoblastu, tedy vyživovací tkáni, jejíž všeobecně známou částí je placenta. Pro svůj vzhled je tento patologický trofoblast označován jako úplná zásněť hroznová neboli mola hydatidosa completa, kompletní mola. Žádné embyo se nevytváří, zato ale v děloze ženy rychle narůstá tkáň připomínající hrozny révy, tvořená měchýřky vodnatých klků. Ve státech s dobrou péčí o těhotnou je patologie brzy rozpoznána, ale tam, kde tato péče chybí, může mola dosáhnout značných rozměrů. Kromě rizika krvácivých stavů v sobě skrývá ještě další nebezpečí: Tkáň se může zvrhnout až v invazívní, metastázující choriokarcinom.
„Že se nejedná o jalové dovádění na poli pusté teorie, to dokládá autorčin úplně první forenzní případ, který v červenci 2002 přistál na jejím laboratorním stole: jednalo se o pohlavní zneužívání čtrnáctileté dívky, při němž byla počata kompletní dispermická mola.“
Ještě v 80. letech 20. století najdeme v odborných článcích autorská podivení nad tím, proč se diandrická diploidie od té normální tak zásadně liší. Dnes již víme, že hlavní příčinou je savčí genomický imprinting – zvláštní mechanismus regulace genové exprese. U naprosté většiny našich autozomálních genů buňka exprimuje (= překládá do struktury bílkovin) obě kopie daného genu, tedy tu z mateřského i z otcovského chromozómu, u pár set genů to neplatí: jsou totiž imprintované. To znamená, že je pomocí metylace trvale umlčena buď mateřská, nebo otcovská kopie genu,3) a užívána je ta druhá. O důvodech tohoto evolucí přijatého „aktivního opatření“ se stále diskutuje, jedna z hlavních hypotéz jej však spojuje s tlumením konfliktu mateřských a otcovských zájmů během nitroděložního vývoje: zatímco otcovým zájmem je pod heslem „z cizího krev neteče“ maximalizovat výživu plodu i za cenu značného vyčerpání mateřského organismu, zájmem matky je dobrý rozvoj embrya s umírněným růstem a schopností efektivního využití omezenějšího množství živin. Tomu odpovídá i povaha a chování kompletní moly, tvořené výhradně genetickým materiálem paternálního původu: agresivní, silně rozvinutá vyživovací tkáň bez založeného embrya.
Příliš mnoho chromozomálních sad
Vyhráno ale není, ani když je vajíčko normální, tedy haploidní. Během oplození vajíčko obvykle nejprve komunikuje s mnoha spermiemi, z nichž pouze jediná nakonec dostane kýžené vízum, zatímco ostatním je vstup odepřen. Tento mechanismus však může vzácně selhat, a do vajíčka se tak podaří proniknout spermiím dvěma (opět mluvíme o dispermii), v důsledku čehož vzniká biparentální monogynický diandrický triploid. I u něj pozorujeme důsledky nevyváženého poměru maternálního a paternálního komplementu: embryo se sice zakládá, ale s četnými těžkými vývojovými vadami, a některé klky trofoblastu mají molární charakter. Proto tuto entitu označujeme jako mola hydatidosa partialis - částečná mola. Ta může vzniknout ještě jiným mechanismem: tak, že normální vajíčko je oplozeno diploidní spermií.

Diploidní může být ovšem v důsledku chyby při oogenezi i samo vajíčko. Po vstupu jediné normální spermie vzniká také triploidní buňka, tentokráte je ale nepoměr v množství maternálních a paternálních chromozómů obrácený – jedná se o biparentálního digynického monoandrického triploida. I toto embryo má řadu charakteristických vývojových vad, trofoblast je ale celkově méně rozvinutý a nemá molární povahu; mluvíme proto o nonmolární triploidii. Za další vzácný, nicméně možný mechanismus jejího vzniku je považována tzv. diovie, při níž dojde ke splynutí dvou normálních vajíček a jejich následnému oplození normální spermií. Nonmolární triploidie může být někdy slučitelná s krátkodobým přežitím: v literatuře je popsán případ dítěte s touto poruchou, které se dožilo šesti měsíců. Protože ale laskavému čtenářstvu už jistě jde hlava kolem, odskočme si nyní na chvilku do světa nás běžných, nudných normoploidů.

Je Karel Honzíkovým otcem?
Vztah Karla Mráčka s Lucií Slunečnou se odvíjel přesně podle páně Mráčkových představ, ale jen do té doby, než mu slečna Slunečná oznámila, že je s ním v očekávání, a než za dalšího půl roku povila krásného, zdravého synka Honzíka. Dílem proto, že založení rodiny zatím jaksi neměl v úmyslu, hlavně ale proto, že přeochotní vrabci z firmy mu brzy vycvrlikali, že nebyl sám, kdo půvabnou sekretářku brával čas od času na večeři. Také Hromek z obchodního a Měsíček z distribuce u ní nepřišli zkrátka. Znejistěný Karel Mráček proto usoudil, že za vola být nechce, a své otcovství vůči Honzíkovi popřel. Protože však Lucie Slunečná trvala na tom, že Mráček otcem je, doputoval jejich spor – jako mnoho jiných takových – až k soudu, který nařídil provedení paternitního testu.
Určení otcovství analýzou DNA je založeno na porovnání genotypu dítěte, matky a domnělého otce. Za normálních okolností přitom vycházíme z jednoduchého předpokladu: genotyp libovolného autozomálního lokusu dítěte vzniká složením ze dvou rodičovských alel, a to tak, že dítě dostane do vínku jednu náhodně vybranou alelu matky a jednu náhodně vybranou alelu otce. V našem případě byl v testovaném lokusu u Lucie Slunečné zjištěn genotyp 12/17, u Karla Mráčka 15/18 a u Honzíka 12/15. Není-li o Luciině mateřství vůči Honzíkovi pochyb, pak Honzíkova alela 12 je téměř jistě mateřská, a alela 15 tudíž musí být otcovská.
Další postup vážení tohoto důkazu je založen na principech tzv. bayesovské inference, která pracuje s podmíněnými pravděpodobnostmi jevů a o níž si lze zevrubně přečíst v článku Detektivka ze Stínadel. Na stole před znalcem leží dvě možné hypotézy: první, že shoda v alele 15 mezi Honzíkem a Karlem Mráčkem nastala kauzálně, neboť Karel je Honzíkův biologický otec, a druhá, že tato shoda nastala náhodou a Honzíkovým otcem je jiný nepříbuzný muž z téže populace. Znalec stanoví, jak pravděpodobná je pozorovaná shoda za platnosti jedné a druhé hypotézy; poměr těchto pravděpodobností se v případě určování otcovství nazývá paternitní index a je jedinou korektní hodnotou, pomocí níž by znalec měl sílu provedeného důkazu vyjádřit.
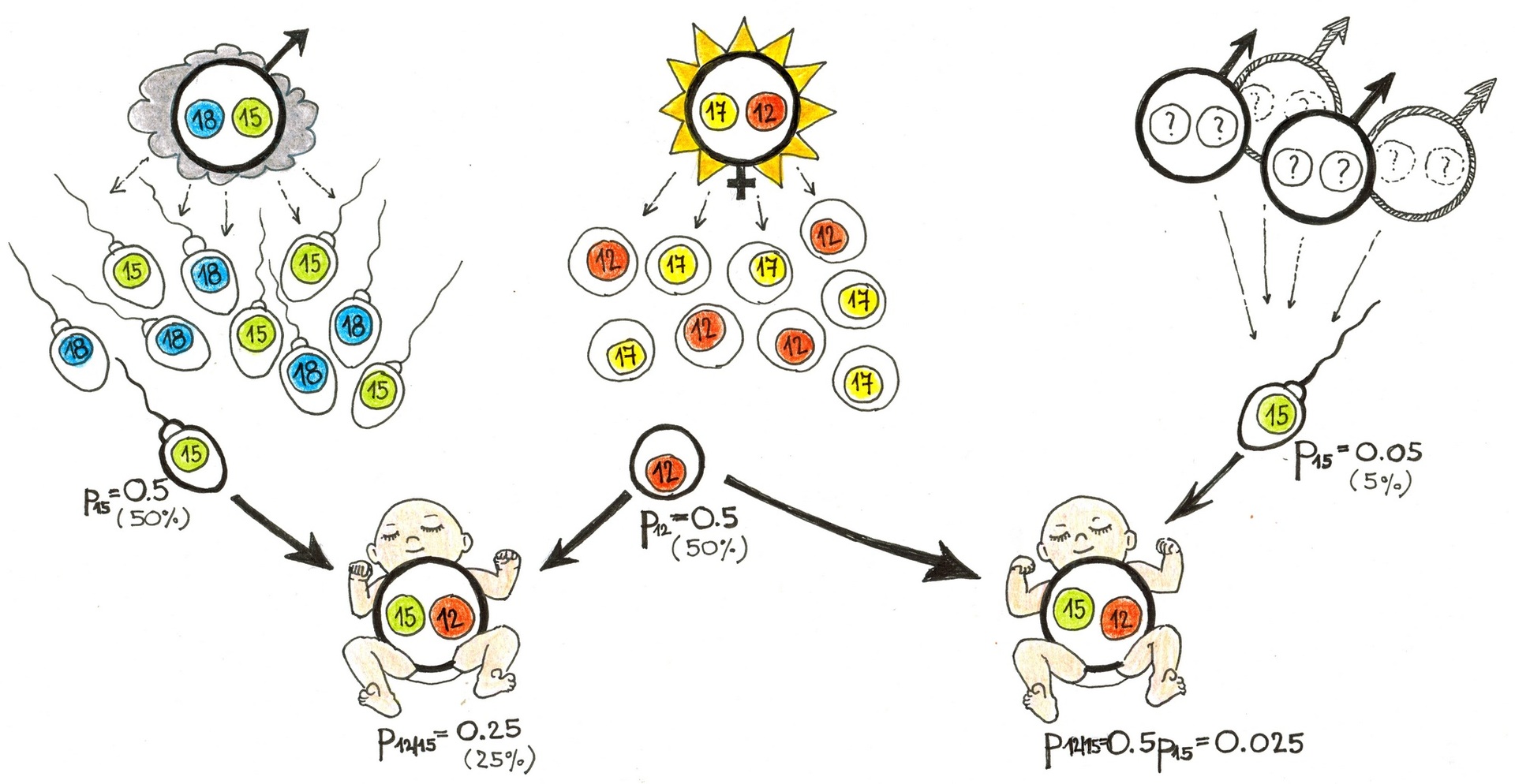
Pokud je Honzíkovým otcem Karel, pak pravděpodobnost, že Lucie Honzíkovi ze svého genotypu 12/17 předá právě alelu 12 a současně mu Karel ze svého genotypu 15/18 předá právě alelu 15, je 1/2 × 1/2, neboli 25 %. Pokud je naopak Honzíkovým otcem jiný nepříbuzný muž, pak pravděpodobnost, že Lucie Honzíkovi ze svého genotypu 12/17 předá právě alelu 12 a současně mu neznámý muž předá právě alelu 15, je 1/2 × p15, kde p15 je četnost alely 15 v populaci. Dejme tomu, že ta je 5 % neboli 0,05. Paternitní index pak spočítáme snadno jako 0,25/0,025 = 10.
Toto číslo nám říká, že danou konstelaci genotypů Lucie, Karla a Honzíka budeme pozorovat desetkrát pravděpodobněji, pokud Karel otcem je, než pokud jím není, jinak řečeno, že ať jsou ostatní důkazy o otcovství Karla vůči Honzíkovi jakkoliv silné, toto zkoumání je posiluje desetkrát. Testování jednotlivého lokusu by samozřejmě bylo nedostatečné – v praxi se jich analyzuje cca 15–30, takže výsledný paternitní index může dosáhnout třeba hodnoty řádově 1015–1020. To už představuje extrémně vysokou míru jistoty, že daný muž je skutečně biologickým otcem.
čtěte také:
Divoký tanec pravděpodobností
Jak ale přistoupit k paternitním (a maternitním) testům tehdy, pokud výsledkem početí není normoploidní Honzík, ale některý z výše popsaných hrubě aberantních produktů početí? Zavedená pravidla kombinování alel tu neplatí, a tak si pro výpočet indexů musíme pro každý jednotlivý typ patologie odvodit pravidla nová. Ukažme si to třeba na diandrické triploidii, vzniklé dispermií, tedy když do normálního vajíčka pronikly dvě normální spermie téhož muže.4)
Dejme tomu, že matka má genotyp 12/13, nařčený muž 13/14 a parciální mola 12/13/14, přičemž alela 12 má v populaci frekvenci 5 %, alela 13 frekvenci 15 % a alela 14 frekvenci 10 %. Jak už jsme si ukázali výše, paternitní index kostruujeme tak, že proti sobě postavíme dvě alternativní hypotézy – H1: nařčený muž je biologickým původcem moly, a H2: jiný nepříbuzný muž je biologickým původcem moly.

Nejprve se budeme na zjištěné genotypy dívat optikou první hypotézy – zajímá nás tedy, jak pravděpodobné s ohledem na genotyp matky a genotyp nařčeného muže je, že jimi zplozená dispermická parciální mola bude mít právě ten genotyp, který má. Víme, že jedna alela v genotypu moly je matčina, zbylé dvě tedy musejí být otcovy. V našem případě existuje jediná možnost, jak se mohly alely matky a nařčeného muže kombinovat: matčino vajíčko muselo nést alelu 12, jedna mužova spermie alelu 13 a druhá mužova spermie alelu 14. Jak pravděpodobné je, že se to stane? Pravděpodobnost, že vajíčko ponese alelu 12, je 0,5 neboli 50 %. Protože první náhodně vybraná spermie může být 13 a druhá 14, nebo obráceně, pravděpodobnost, že dvě náhodně vybrané spermie ponesou alely 13 a 14, je (1/2 × 1/2) + (1/2 × 1/2) = 0,5 neboli 50 %. Celková pravděpodobnost vzniku genotypu 12/13/14 je tedy při platnosti H1 rovna 0,5 × 0,5 = 0,25 neboli 25 %.
Teď si naopak představíme, že platí H2, tedy že rodiči parciální dispermické moly s genotypem 12/13/14 jsou testovaná žena s genotypem 12/13 a neznámý muž. Možnosti, jak mohla mola vzniknout, už budou v tomto případě dvě: buďto do vajíčka s alelou 12 pronikla jedna spermie s alelou 13 a jedna spermie s alelou 14 (neznámý muž tedy musel mít genotyp 13/14), nebo do vajíčka s alelou 13 pronikla jedna spermie s alelou 12 a jedna spermie s alelou 14 (neznámý muž tedy musel mít genotyp 12/14). Nezbývá než si pěkně „zakombinatorikovat“: pravděpodobnost, že vajíčko testované ženy nese alelu 12, je 50 %, pravděpodobnost, že náhodný muž má genotyp 13/14, je 2p13p14, kde p13 a p14 jsou populační frekvence těchto alel (zadali jsme si je výše, 15 % a 10 %) – dosazením získáme pravděpodobnost genotypu 13/14 rovnu 3 % a pravděpodobnost, že jedna spermie bude 13 a druhá 14, je 50 %, celkově tedy 0,5 × 0,03 × 0,5 = 0,0075 neboli 0,75 %. Teď druhá možnost: pravděpodobnost vajíčka s alelou 13 u testované ženy je 50 %, pravděpodobnost, že náhodný muž má genotyp 12/14, je 2p12p14, což po dosazení příslušných populačních frekvencí dá 1 %, a pravděpodobnost, že jedna spermie bude 12 a druhá 14, je 50 %, celkově tedy 0,5 × 0,01 × 0,5 = 0,0025 neboli 0,25 %. Výsledná pravděpodobnost obou možností je 0,75 % + 0,25 %, tedy 1 %.
Paternitní index už má pak standardní konstrukci: je to podíl první pravděpodobnosti (tj. při platnosti hypotézy, že nařčený muž je otcem) a druhé pravděpodobnosti (tj. při platnosti hypotézy, že neznámý muž je otcem). V našem případě to bude krásných 0,25/0,01 = 25.
Kdo by se snad cítil při všem tom počítání poněkud ztracen, nechť věří, že může být i hůř: u některých aberantních produktů početí je při určitých konstelacích genotypů ještě mnohem více možných variant kombinování alel prostřednictvím vajíček a spermií. Do hry také mohou vstoupit další faktory – kupříkladu k diploidii vajíčka či spermie může dojít z více příčin, a každá z nich vede k jiné pravděpodobnosti, že genotyp původce bude v gametě tvořen oběma jeho alelami (tedy například, že diploidní spermie muže s genotypem 15/17 ponese také genotyp 15/17, a ne 15/15 nebo 17/17). Model je pak nutno rozšířit o další zdroje nejistoty. Svět aberantních produktů početí se tak pro forenzního genetika či genetičku stává fascinujícím rejdištěm, kde lze popustit uzdu své fantazii a dosyta se bayesovsky vyřádit.
A že se nejedná o jalové dovádění na poli pusté teorie, to dokládá autorčin úplně první forenzní případ, který v červenci 2002 přistál na jejím laboratorním stole: jednalo se o pohlavní zneužívání čtrnáctileté dívky, při němž byla počata kompletní dispermická mola. Vskutku znalecký křest ohněm.
Poznámky
1) Biparentální = 2 rodiče; monogynický = 1 sada chromozómů od ženy; monoandrický = 1 sada chromozómů od muže; diploid =2 sady chromozómů celkem.
2) Příčiny jsou dvě: za prvé, Y chromozóm je extrémně „genově chudý“, takže případná změna genové dávky se týká jen nepatrného počtu genů v buňce, a za druhé, X chromozóm disponuje mechanismem zvaným lyonizace, kterým se v buňkách ženy jeden ze dvojice X chromozómů „svine“ a tím trvale inaktivuje; tento proces pak může proběhnout i v buňkách s vyšším počtem X chromozómů, kde lyonizují všechny až na jeden.
3) Komu konvenují terminologické hrátky, užije si i tady: takový gen pak totiž podle povahy imprintingu označujeme jako maternálně imprintovaný paternálně exprimovaný, nebo paternálně imprintovaný maternálně exprimovaný.
4) Mimochodem, u dispermie si lze představit ještě větší forenzně genetický bizár: dvě spermie proniklé do téhož vajíčka totiž nemusejí nutně být od jednoho muže. Pokud se do pohlavních orgánů ženy dostane v krátkém časovém rozmezí ejakulát dvou mužů (možné mechanismy ponecháváme na fantazii p. t. čtenářstva), může být vajíčko teoreticky oplodněno i spermiemi dvěma různých mužů, a výsledkem je triparentální monogynický diandrický triploid. Protože ale podmínkou jeho vzniku je synchronizace dvou dějů, které jsou vzácné samy o sobě (k tomu podotýkáme, že pornografické materiály nejsou zdrojem relevantních informací o většinovém chování populace), bude tato genetická entita extrémně raritní.
O autorovi
Halina Šimková

Další články k tématu
Detektivka ze Stínadel
Čichám, čichám zločin
DNA: důkaz mocný, nikoli všemocný
Zrození a dospívání forenzní genetiky
Kognitivní zkreslení v kriminalistice
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů
