









Rostliny C4
Tímto akronymickým a pro mnohé jistě i tajemným pojmem označují fyziologové skupinu rostlin, jejichž příjem CO2 a mnohé další vlastnosti se liší od většiny ostatních rostlin. Proč vlastně metabolismus C4 vznikl? Jaké důsledky přináší svým nositelům a celým společenstvům?
 Pojem fotosyntéza vyvolává u mnoha lidí představu procesu, ve kterém vzniká kyslík. Přestože typů fotosyntézy je mnoho, ta oxygenní je dnes na Zemi nejrozšířenější a pro naši existenci zcela zásadní. Rostliny při fotosyntéze pohlcují světlo (přesněji záření v rozsahu vlnových délek asi 400–700 nm) a s pomocí jeho energie zabudovávají anorganický oxid uhličitý (CO2) do organických složek svého těla. Fotosyntéza rostlin je složitý a několikastupňový proces. Zaměřme se pouze na části související s příjmem oxidu uhličitého, které nejvíce odrážejí právě odlišnosti rostlin C4.
Pojem fotosyntéza vyvolává u mnoha lidí představu procesu, ve kterém vzniká kyslík. Přestože typů fotosyntézy je mnoho, ta oxygenní je dnes na Zemi nejrozšířenější a pro naši existenci zcela zásadní. Rostliny při fotosyntéze pohlcují světlo (přesněji záření v rozsahu vlnových délek asi 400–700 nm) a s pomocí jeho energie zabudovávají anorganický oxid uhličitý (CO2) do organických složek svého těla. Fotosyntéza rostlin je složitý a několikastupňový proces. Zaměřme se pouze na části související s příjmem oxidu uhličitého, které nejvíce odrážejí právě odlišnosti rostlin C4.
U „běžných“ rostlin, řas a sinic je CO2 biologicky vázán prostřednictvím enzymu ribulosa-1,5-bisfosfátkarboxyláza/oxygenáza neboli Rubisco. Prvním stabilním produktem fotosyntézy u těchto rostlin je tříuhlíkatá látka, a proto jim fyziologové říkají C3. Rubisco je evolučně velmi starý enzym a v době jeho vzniku panovaly zcela jiné podmínky než dnes. Především bylo v atmosféře mnohonásobně víc CO2. Proto se neprojevovala nevýhoda dvojfunkčnosti enzymu, která je zřejmá z jeho názvu a která jeho účinnost v dnešních podmínkách výrazně omezuje. Enzym totiž kromě CO2 (karboxylace) váže i kyslík (oxygenace) – tím více, čím více kyslíku a méně oxidu uhličitého je v jeho okolí přítomno. Navázáním O2 však vzniká „špatný produkt“ – glykolát – a uhlík v něm je zachráněn jen ze dvou třetin v energeticky náročném procesu nazývaném fotorespirace. Poslední třetina je odbourána až na oxid uhličitý, který je vydán zpět do mezibuněčného vzduchu.
 Fotorespirace je složitou kaskádou reakcí – uhlík v glykolátu prochází řadou přeměn, až se z části (2/3) stává opět tříuhlíkatá látka – regulérní produkt asimilace CO2. Zajímavé je, že k „úspěšnému zachraňovacímu procesu uhlíku“, kterým fotorespirace je, je třeba spolupráce tří buněčných organel – chloroplastu, mitochondrie a peroxizomu. Fotorespirace se tedy navenek podobá dýchání, které je však vázáno jen na světlo (a běžící fotosyntézu). Odtud jeho jméno.
Fotorespirace je složitou kaskádou reakcí – uhlík v glykolátu prochází řadou přeměn, až se z části (2/3) stává opět tříuhlíkatá látka – regulérní produkt asimilace CO2. Zajímavé je, že k „úspěšnému zachraňovacímu procesu uhlíku“, kterým fotorespirace je, je třeba spolupráce tří buněčných organel – chloroplastu, mitochondrie a peroxizomu. Fotorespirace se tedy navenek podobá dýchání, které je však vázáno jen na světlo (a běžící fotosyntézu). Odtud jeho jméno.
Záměnnost kyslíku a oxidu uhličitého u Rubisco zákonitě nebyla na překážku v době vzniku fotosyntézy, v prahorách, kdy CO2 tvořil hlavní složku atmosféry. Oxygenace byla zanedbatelná i v době, kdy byla koncentrace CO2 několikanásobná, než je dnes. Rubisco totiž mnohem ochotněji váže CO2 než kyslík (říkáme, že má k CO2 větší afinitu). Navíc se CO2 ve vodě (buňkách) rozpouští asi 50× lépe než kyslík.
 Takřka kritickou se situace stala až koncem třetihor (před několika miliony let) a hlavně ve čtvrtohorách. V této době už byla většina oxidu uhličitého ze vzduchu vázána ve vápencích, uhlí, ropě a živých i odumřelých tělech organismů a fotosyntézou zaměněna ve vzduchu za kyslík. Připomeňme, že tento kyslík nevzniká v rostlinách štěpením CO2, nýbrž vody. Nejméně CO2, pokud dnes víme, bylo ve vzduchu během dob ledových, přibližně pouhé 0,02 % objemu. Naopak v dobách meziledových jeho obsah vzrůstal asi o třetinu (patrně hlavně díky uvolňování CO2 rozpuštěného v oceánech). Dnes je ve vzduchu CO2 trochu více – 0,039 % objemu – a jak známo dále stoupá. Pro rostliny je to však stále málo, uvážíme-li, že koncentrace kyslíku ve volné atmosféře je skoro 600× vyšší než oxidu uhličitého (21 % oproti 0,039 %). Při této situaci jednak není Rubisco nasyceno (nepracuje svou maximální rychlostí), jednak fotorespirací prochází kolem 20–40 % uhlíku získaného fotosyntézou, který se dostal do špatného produktu – glykolátu. To představuje energetické plýtvání a část CO2 se vrací zpět do atmosféry. Rychlost oxygenace (a tudíž i ztrátové fotorespirace) navíc vzrůstá s teplotou. Souvisí to jak s vlastnostmi enzymu Rubisco, tak s rozpustností kyslíku a oxidu uhličitého ve vodě (buněčných tekutinách).
Takřka kritickou se situace stala až koncem třetihor (před několika miliony let) a hlavně ve čtvrtohorách. V této době už byla většina oxidu uhličitého ze vzduchu vázána ve vápencích, uhlí, ropě a živých i odumřelých tělech organismů a fotosyntézou zaměněna ve vzduchu za kyslík. Připomeňme, že tento kyslík nevzniká v rostlinách štěpením CO2, nýbrž vody. Nejméně CO2, pokud dnes víme, bylo ve vzduchu během dob ledových, přibližně pouhé 0,02 % objemu. Naopak v dobách meziledových jeho obsah vzrůstal asi o třetinu (patrně hlavně díky uvolňování CO2 rozpuštěného v oceánech). Dnes je ve vzduchu CO2 trochu více – 0,039 % objemu – a jak známo dále stoupá. Pro rostliny je to však stále málo, uvážíme-li, že koncentrace kyslíku ve volné atmosféře je skoro 600× vyšší než oxidu uhličitého (21 % oproti 0,039 %). Při této situaci jednak není Rubisco nasyceno (nepracuje svou maximální rychlostí), jednak fotorespirací prochází kolem 20–40 % uhlíku získaného fotosyntézou, který se dostal do špatného produktu – glykolátu. To představuje energetické plýtvání a část CO2 se vrací zpět do atmosféry. Rychlost oxygenace (a tudíž i ztrátové fotorespirace) navíc vzrůstá s teplotou. Souvisí to jak s vlastnostmi enzymu Rubisco, tak s rozpustností kyslíku a oxidu uhličitého ve vodě (buněčných tekutinách).
 Fotorespirace však má pro rostlinu i svůj přínos. Stala se dnes důležitým odpadním kanálem pro přebytečnou světelnou energii. Tu sice v podmínkách nedostatku CO2 proměňuje zčásti v neužitečné teplo, ale tím zároveň chrání fotosyntetický aparát před poškozením. Krom toho má svoji úlohu v dusíkovém metabolismu.
Fotorespirace však má pro rostlinu i svůj přínos. Stala se dnes důležitým odpadním kanálem pro přebytečnou světelnou energii. Tu sice v podmínkách nedostatku CO2 proměňuje zčásti v neužitečné teplo, ale tím zároveň chrání fotosyntetický aparát před poškozením. Krom toho má svoji úlohu v dusíkovém metabolismu.
Vynález C4
Záměnnost kyslíku za oxid uhličitý u enzymu Rubisco začala rostliny ve velké míře omezovat zřejmě v miocénu, středních třetihorách, tedy asi před 10 miliony lety. Tenkrát poprvé (pokud dnes víme) klesla koncentrace CO2 pod 0,05 % objemu vzduchu a oxygenace tak dosáhla téměř současných hodnot. Jak se rostlinám C4 podařilo handicap v podobě fotorespirace zmírnit?
 Možností by bylo více. Co třeba nahradit Rubisco enzymem obdařeným jen karboxylační aktivitou, tedy vážícím jen CO2? Taková záměna nebo vylepšení klíčového enzymu se však přírodě nikdy nepovedla. Navíc, ač se to třeba nezdá, času na to bylo velmi málo – co je desítka milionů let problémů proti třem miliardám, kdy to fungovalo dobře. Řešení se přesto našlo, nicméně poněkud kompromisní.
Možností by bylo více. Co třeba nahradit Rubisco enzymem obdařeným jen karboxylační aktivitou, tedy vážícím jen CO2? Taková záměna nebo vylepšení klíčového enzymu se však přírodě nikdy nepovedla. Navíc, ač se to třeba nezdá, času na to bylo velmi málo – co je desítka milionů let problémů proti třem miliardám, kdy to fungovalo dobře. Řešení se přesto našlo, nicméně poněkud kompromisní.
Když se snížila koncentrace CO2 v atmosféře Země, nešlo by alespoň zvýšit koncentraci CO2 v místě, kde se nachází Rubisco? Tím by se oxygenace částečně nebo skoro úplně potlačila. Vytváření koncentračních gradientů je pro živé tvory typické – je to vlastně hlavní funkce biologických membrán a přenašečů v nich. U mnohých skupin řas a sinic nacházíme celou řadu pump, které pumpují oxid uhličitý přes membrány a koncentrují ho uvnitř daného prostoru. Nás však bude zajímat ta, která vedla ke vzniku suchozemských C4 rostlin.
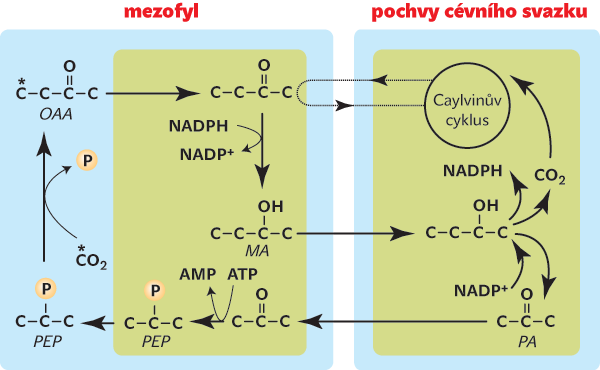
Jde o spolupráci dvou anatomicky a biochemicky odlišných typů buněk. Ty, které obklopují cévní svazky (buňky pochev cévních svazků), obsahují Rubisco a přijímají CO2 jako ostatní rostliny. Buňky vnější (mezofylové) zajišťují příjem atmosférického CO2 a předávají ho ve vázané podobě buňkám poševním. V nich je molekula CO2 asimilována Rubisco. Důležité je, že mezofylové buňky váží CO2 jiným enzymem, který nejeví oxygenační aktivitu, je rychlejší a navíc má k CO2 vyšší příchylnost. Jmenuje se fosfoenolpyruvátkarboxyláza (PEPc), protože váže CO2 na tříuhlíkatý fosfoenolpyruvát, čímž vzniká čtyřuhlíkatý oxalacetát (obr. 2). Podle počtu uhlíků v oxalacetátu vznikl název metabolismus C4.


 PEPc je podobně jako Rubisco velmi starý enzym, který plní různé funkce, a to nejen v rostlinách. Je například nepostradatelný při otevírání průduchů a k vytváření metabolické rovnováhy mezi organelami (chloroplasty a mitochondriemi) – poskytuje uhlík v podobě kyseliny jablečné pro mitochondriální metabolismus. V rostlinách C4 se navíc ukázala jeho vhodnost pro příjem CO2 ze vzduchu v případě, že v něm je CO2 málo. Rostliny C4 tedy ke svému vzniku nepotřebovaly vlastně téměř nic nového. Snad proto se v geologickém čase objevují celkem náhle.
PEPc je podobně jako Rubisco velmi starý enzym, který plní různé funkce, a to nejen v rostlinách. Je například nepostradatelný při otevírání průduchů a k vytváření metabolické rovnováhy mezi organelami (chloroplasty a mitochondriemi) – poskytuje uhlík v podobě kyseliny jablečné pro mitochondriální metabolismus. V rostlinách C4 se navíc ukázala jeho vhodnost pro příjem CO2 ze vzduchu v případě, že v něm je CO2 málo. Rostliny C4 tedy ke svému vzniku nepotřebovaly vlastně téměř nic nového. Snad proto se v geologickém čase objevují celkem náhle.
Evoluce syndromu C4
Současná představa vývoje syndromu C4 se opírá zejména o pozorování druhů, jež vykročily směrem k břehům C4, kterých ovšem ještě nedosáhly (např. rod Flaveria z čeledi hvězdnicovitých). Zbývá jim ještě různě dlouhá cesta ukazující evoluci v akci. Jeden scénář vzniku syndromu C4 je tento:
1. Zdvojí se řada genů, což je (nejen) u rostlin běžné. Jedna kopie plní svůj účel, druhá může mutovat k nové nebo pozměněné funkci, která se může časem hodit.
2. Zhoustne žilnatina listů a posléze se vytvoří věnčitá anatomie. Zvýší se tím podíl cév vedoucích vodu do listu. Dělají to i C3 druhy suchých oblastí.
3. Zvýší se množství chloroplastů v pochvách cévních svazků (jen u trav). Trávy C3 v nich jinak skoro žádné nemají (žilky jsou průsvitné). Jsou-li však žilky hodně hustě, hodně světla listem prosvítá, což snižuje fotosyntézu, a tím i konkurenční sílu druhu.
4. Posílí se transport glykolátu do pochev cévních svazků. Fotorespirace v mezofylu je tím omezena, což zlepší příjem CO2 ze vzduchu při jeho nižších koncentracích. Toto je typická vlastnost mnohých dnešních druhů přechodných mezi C3 a C4.
5. Zvýší se aktivita PEPc v mezofylu i dalších enzymů. Výsledkem bude „pumpování“ vázaného CO2 do buněk cévních pochev.
6. Vznikne kontakt mezi mezofylovými buňkami a buňkami cévních pochev, které budou spojeny četnými plazmodezmy a zároveň izolované korkem, aby plynný CO2 neutíkal.
7. Systém se „doladí“. Např. se potlačí Rubisco v mezofylových buňkách, kde se stává zbytečným, protože PEPc váže CO2 účinněji.
Tento pohled je notně zjednodušený a navíc jen jeden z možných. Za povšimnutí stojí, že časnější kroky nesou rostlině výhodu samy o sobě i bez vyhlídky vzniku fotosyntézy C4 a pozdější fáze jsou zase nutnými nebo pravděpodobnými důsledky těch dřívějších. Vývoj pumpy C4 – a to je důležité – tedy mohl být postupný. Nejprve se objevil sklon k recyklování CO2 z fotorespirace. Postupně se pumpa vylepšovala, až u vyšších rostlin vedla k vzniku druhů C4, jak je známe dnes. Přitom Rubisco pracovalo nerušeně dále, jen mu bylo postupně „přilepšováno“.
Nic není zadarmo
 Proč tedy mechanismus C4 neprovozují všechny rostliny? Proč ty, které to umí, nevytlačí druhy C3? Proč dnes nejsou jen rostliny C4?
Proč tedy mechanismus C4 neprovozují všechny rostliny? Proč ty, které to umí, nevytlačí druhy C3? Proč dnes nejsou jen rostliny C4?
Jednoduchá odpověď: Protože pumpa C4 řeší kauzu Rubisco jen částečně. Složitější odpověď: Kdyby nebyla oxygenace, rostliny C3 by potřebovaly k celkem navázání jedné molekuly CO2 3 molekuly ATP (univerzálního energetického platidla buňky) a 2 molekuly NADPH (reduktant). Rostliny C4 to samé a navíc 2 ATP na zajištění funkce pumpy CO2, dohromady 5 ATP. Zdálo by se tedy, že rostliny C4 jsou vždy v nevýhodě, protože méně účinně využívají světlo (s pomocí jehož energie se vytváří ATP i NADPH). Hodnota ATP pro druhy C3 je však v přírodě vždy vyšší právě kvůli vázání kyslíku a složitému zachraňování části látek takto vzniklých. Při podílu fotorespirace asi 30 % fotosyntézy je to právě 5 ATP jako u rostlin C4. Ve výsledku se tedy metabolismus C4 vyplácí v případě, že fotorespirace rostlin C3 činí asi třetinu fotosyntézy a více. V opačném případě (fotorespirace menší než 1/3 čisté fotosyntézy) jsou náklady na provoz pumpy C4 větší než zisk, který přinese. (Jakým podmínkám tato situace odpovídá v přírodě a kde druhy C4 přednostně rostou, si řekneme v dalším díle.)
Kdy a u kterých skupin se fotosyntéza C4 objevuje?
Výzkum DNA ukázal, že rostliny C4 vznikly vícekrát nezávisle. Zatímco v knize C4 Plant Biology z r. 1999 se píše o nejméně 31 počátcích, Rowan Sage ve svém článku o evoluci rostlin C4 z r. 2004 uvádí již 45 až 48 nezávislých vzniků! Výzkum běží rychle kupředu a výsledky jistě nejsou definitivní. Co se však už dnes jeví zřejmé, je jakási náchylnost rostlin C3 stát se C4.
Čím je věda „dál“, tím soudobějšími rostliny C4 shledává. Extrémní odhady jsou např. prvohorní karbon (asi před 300 miliony let) – v době, ve které (jak dnes víme) nebyly ještě kvetoucí rostliny. Ještě r. 1999 najdeme mutační výpočet stáří trav C4 75 milionů let. Nejnovější práce, která se souborně věnuje vzniku a vývoji rostlin C4, však hovoří o „pouhých“ 20–35 milionech let (u trav). Pro srovnání, nejstarší fosilie se zachovanou věnčitou anatomií (znak současných trav C4) je stará 12,5 milionu let. Od této doby můžeme jejich existenci snad považovat za prokázanou.
Dvouděložné druhy C4 jsou však pravděpodobně ještě mladší a zdá se, že vznikají dodnes. Tomu nasvědčuje i skutečnost, že zatímco celé rody travin jsou většinou buď C3 nebo C4, u dvouděložných je to často v rámci jednoho rodu „namíchané“. Existují však, jak to tak bývá, výjimky. Např. dvouděložné laskavce jsou všechny C4, naproti tomu tráva Alloteropsis semialata, původem z Afriky, může být C3 i C4. Krásným příkladem evolučního mládí dvouděložných je již zmíněný rod Flaveria. V něm najdeme jak zástupce C3 a C4, tak druhy, které jsou „na půli cesty“.
Počet dnes známých druhů C4 se blíží 10 000. Více než polovinu z nich tvoří trávy (lipnicovité – Poaceae, viz tab. I). (Eko)fyziologické vysvětlení jejich úspěšnosti však není
tak jednoduché. Evoluční zjednodušení je následující: Metabolismus C4 vznikl u trav dříve než u ostatních skupin. Proto měl víc času „vyladit se“, aby poskytoval maximální výhody. Navíc být trávou je samo o sobě celosvětově extrémně úspěšná strategie bez ohledu na typ metabolismu. Opět není zcela jasné proč. U dvouděložných na druhou stranu překvapuje počet čeledí, ve kterých se syndrom C4 objevil a objevuje.
Kdy si vegetace C4 podmanila Zemi?
Jako se fotosyntéza C3 a C4 výrazně liší ve svém principu, liší se i ve volbě oxidu uhličitého s lehkým (12C) a těžkým (13C) izotopem uhlíku. Výsledkem je jejich odlišné zastoupení v biomase rostlin C3 a C4, ale také ve fosiliích býložravců, kteří se jimi živili. K obrovské izotopové změně v zubní sklovině kopytníků a ve fosilizovaných skořápkách vajec pštrosů došlo na mnoha (tehdy) teplých místech světa před 8–5 miliony let. Tento zlom se obecně považuje za vítězství C4 druhů v teplých travinných biotopech. Koncentrace CO2 se však v oné době téměř nelišila od té dnešní. Před 15–5 miliony let se vzdušná koncentrace CO2 navíc nijak dramaticky nesnižovala. Hlavní roli v postupném šíření vegetace C4 tedy patrně sehrálo klima, které se postupně stávalo v mnoha částech tropů, subtropů a teplejšího mírného pásu sušší a sušší.
Největšího zvýhodnění se rostliny s pumpou CO2 dočkaly během dob ledových. Přestože u nás převažovala tundra, blíže k rovníku se ochladilo jen málo. Navíc bylo mnohde sušší podnebí, což přispělo k dalšímu rozmachu druhů C4 na šířícím se bezlesí. Přitom nešlo jen o šíření. Silný evoluční tlak (soudobá výhodnost „být C4“) nepochybně urychlil i vznik a vývoj nejmladších skupin C4, hlavně dvouděložných.
Svět před fenoménem C4 a s ním
Protože i vědci rádi fantazírují, objevují se stále nové spekulace o tom, co rostliny C4 zásadně změnily. Popularizujme zde hypotézu nejodvážnější (z antropocentrického pohledu).
Možná jste si již uvědomili, že vítězné tažení trav C4, které pokryly většinu tropické a subtropické otevřené krajiny, spadá do počátků evoluce člověka. Většina antropologů míní, že by člověk nevznikl, kdyby jeho předkové nemuseli slézt ze stromů. Tato věc se stala v jižní a východní Africe, kolébce lidstva, právě v pozdním miocénu (před 8–5 miliony let), kdy vegetace C4 poprvé nastoupila masově. Těžko můžeme tvrdit, že odlesnění, které donutilo primáty chodit po zemi, bylo způsobeno šířením těchto rostlin. Lesy by patrně ustoupily i bez nich – suchem. Nicméně krajina, ve které se naši předci po další miliony let pohybovali, byla z velké části C4. To mohlo jejich další vývoj nasměrovat.
Podle zubní skloviny fosilních hominidů (hlavně Australopithecus africanus) z doby před 3–2,5 miliony let se odhaduje, že podíl rostlin C4 v jejich jídelníčku (ať už přímo nebo „prohnaných“ přes býložravce) činil nejméně 25 %. Soustavnější izotopové měření fosilií vývojových stupňů člověka však bohužel nemáme. Tyto vykopávky jsou tak vzácné, že jen málokdy bylo dovoleno brousit jim zuby pro izotopové analýzy.
Většinu evoluce tedy člověka provázely druhy C4, často v zapojené vegetaci. Je pravděpodobné, že nebýt jich, vypadáme a konáme dnes jinak.
V příštím pokračování si povíme něco o ekologii a rozšíření rostlin C4.
Ke stažení
 článek ve formátu pdf [873,8 kB]
článek ve formátu pdf [873,8 kB]
O autorovi
Jiří Kubásek

Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
