










Studené nohy bohyně Gaii
Kde budu žít – pomyslel jsem si, než jsem opět usnul –, když můj byt zaroste ledem?
Z básně Led Michala Ajvaze
 Stephen Pyne ve své knize The Ice: A Journey to Antarctica v roce 1986 píše: „Biologie Antarktidy je takřka úplně biologií mořskou. Kromě několika sněžných řas, kolonizujících tající sněhová pole, nežijí žádné organismy výlučně na pevninském ledu; neexistuje terestrická kryo-ekosféra.“ Biologie Antarktidy (a samozřejmě také Arktidy) v roce 2010 ovšem již takovou dávno není. Dnes onen „pustý“ pevninský led zaměstnává molekulární a evoluční biology, fyziology i biogeochemiky.
Stephen Pyne ve své knize The Ice: A Journey to Antarctica v roce 1986 píše: „Biologie Antarktidy je takřka úplně biologií mořskou. Kromě několika sněžných řas, kolonizujících tající sněhová pole, nežijí žádné organismy výlučně na pevninském ledu; neexistuje terestrická kryo-ekosféra.“ Biologie Antarktidy (a samozřejmě také Arktidy) v roce 2010 ovšem již takovou dávno není. Dnes onen „pustý“ pevninský led zaměstnává molekulární a evoluční biology, fyziology i biogeochemiky.
Pro většinu čtenářů je možná překvapující, že spojení slov „ledovcový“ a „ekosystém“ vůbec nese nějaký smysl. Právě relativní jednoduchost života v trvalém chladu však umožňuje studovat nejzákladnější principy života mikrobů ve volné přírodě. Má to i svůj zeměpisný význam. Polární oblasti zabírají 14 % povrchu Země, přibližně 10 % povrchu pevniny je zaledněno, 90 % objemu oceánů má teplotu nižší než 5 °C, více než 70 % sladké vody na Zemi existuje v podobě ledu a velký podíl půd (přibližně 20 %) představuje permafrost.
 Země byla podle řady důkazů několikrát během neoproterozoika téměř zcela zaledněna a nelze vyloučit, že mikrobiální život mohl ve své historii projít i „studeným hrdlem lahve“, což je podstatné i z evolučního hlediska.
Země byla podle řady důkazů několikrát během neoproterozoika téměř zcela zaledněna a nelze vyloučit, že mikrobiální život mohl ve své historii projít i „studeným hrdlem lahve“, což je podstatné i z evolučního hlediska.
Zmrzlou periferii planety, tyto studené nohy bohyně Gaii, alespoň v místech, kde „stojí na pevné zemi“, bychom vám nyní rádi představili jako nedílnou součást jejího půvabného těla.
Sníh
 Sníh je z biotopů kryosféry známý nejdéle, jak ostatně uznal i Pyne.
Sníh je z biotopů kryosféry známý nejdéle, jak ostatně uznal i Pyne.
Každého, kdo navštívil vysoké hory v létě, mohlo někdy zaujmout červené zbarvení tajících sněhových polí. V minulosti byl tento jev interpretován jako Kristova krev, která má připomenout obyvatelům Alp jejich hříchy, a dnes jej lidé často považují za saharský písek. Bystří pozorovatelé v něm však již v 19. století odhalili drobné, červeně zbarvené organismy.
 Jsou to většinou nárosty zelené bičíkaté řasy Chlamydomonas nivalis, u nichž zelenou barvu chlorofylu překrývá červený ochranný pigment astaxantin. Kromě samotné nízké teploty se život v tomto případě musí potýkat s extrémní intenzitou záření (především jeho UV složky). Přesto se na sněhu vyskytují i druhy řas, které syntézu karotenoidů neumějí. Ty lze najít spíš na zastíněných stanovištích nebo hlouběji ve sněhovém profilu. Viditelné zbarvení sněhu je jen zřídka způsobeno sinicemi a mezi fotosyntetickými organismy v tomto prostředí převládají eukaryotické řasy, přestože v chladných terestrických biotopech to typicky bývá obráceně.
Jsou to většinou nárosty zelené bičíkaté řasy Chlamydomonas nivalis, u nichž zelenou barvu chlorofylu překrývá červený ochranný pigment astaxantin. Kromě samotné nízké teploty se život v tomto případě musí potýkat s extrémní intenzitou záření (především jeho UV složky). Přesto se na sněhu vyskytují i druhy řas, které syntézu karotenoidů neumějí. Ty lze najít spíš na zastíněných stanovištích nebo hlouběji ve sněhovém profilu. Viditelné zbarvení sněhu je jen zřídka způsobeno sinicemi a mezi fotosyntetickými organismy v tomto prostředí převládají eukaryotické řasy, přestože v chladných terestrických biotopech to typicky bývá obráceně.

 Spolu s řasami se na tajících sněhových polích vyskytuje i řada bakterií a kvasinek. Moc zatím nevíme, které jsou tam omylem deponovány větrem a spíše přežívají, než žijí, a které jsou v prostředí doma. Víme ale, že ve sněhové pokrývce bývá zpravidla nouze o dostupné živiny, a heterotrofní aktivita bakterií a kvasinek je důležitá právě pro rozklad organické hmoty a návrat živin zpět do hry. Vzhledem k přítomnosti bakteriálních buněk přímo ve slizových vrstvách na povrchu buněk chlamydomonád lze očekávat, že za to od řas dostávají určitou protislužbu v podobě snadno rozložitelných sacharidů. Ať už jde o harmonickou spolupráci nebo o vzájemné vydírání, pro svou jednoduchost by tato společenstva mohla představovat dobrý model pro studium interakcí uvnitř sezonního mikrobiálního společenstva ve volné přírodě.
Spolu s řasami se na tajících sněhových polích vyskytuje i řada bakterií a kvasinek. Moc zatím nevíme, které jsou tam omylem deponovány větrem a spíše přežívají, než žijí, a které jsou v prostředí doma. Víme ale, že ve sněhové pokrývce bývá zpravidla nouze o dostupné živiny, a heterotrofní aktivita bakterií a kvasinek je důležitá právě pro rozklad organické hmoty a návrat živin zpět do hry. Vzhledem k přítomnosti bakteriálních buněk přímo ve slizových vrstvách na povrchu buněk chlamydomonád lze očekávat, že za to od řas dostávají určitou protislužbu v podobě snadno rozložitelných sacharidů. Ať už jde o harmonickou spolupráci nebo o vzájemné vydírání, pro svou jednoduchost by tato společenstva mohla představovat dobrý model pro studium interakcí uvnitř sezonního mikrobiálního společenstva ve volné přírodě.
Aktivita komunity mikroorganismů vázaná na tající sníh je tísněna časem mezi začátkem tání a jeho koncem, kdy sníh zmizí a nový musí být kolonizován každou sezonu znovu z inokula přežívajícího někde v půdě. Následující biotop má na svůj vývoj nesrovnatelně více času.
Supraglaciální biotop
 Centrem biodiverzity a metabolické aktivity ekosystému na povrchu ledovce je tzv. kryokonit. Název vznikl spojením řeckých slov to kryos – ledový, chladný a he konia – prach. Základ pro kryokonit se na povrch ledovce dostává větrem nebo v menší míře s lavinami z nunataků (nezaledněných vrcholů, obklopených ledovcem). Jsou to zrnka hornin, prach a živé i mrtvé organismy. Díky nižšímu albedu (podílu odraženého a dopadajícího záření), než má okolní led, absorbuje více tepla a protává do povrchu ledovce. Tvoří tak jamky s plochým dnem a svislými stěnami, které jsou do určité úrovně naplněny vodou. Jamky mohou dosahovat hloubky až několika decimetrů a plochy v řádech od milimetrů po metry čtvereční. Mohou představovat 2–5 % povrchu ablační zóny ledovce čili oblasti, kde během letní sezony roztaje sníh na povrchu ledu.
Centrem biodiverzity a metabolické aktivity ekosystému na povrchu ledovce je tzv. kryokonit. Název vznikl spojením řeckých slov to kryos – ledový, chladný a he konia – prach. Základ pro kryokonit se na povrch ledovce dostává větrem nebo v menší míře s lavinami z nunataků (nezaledněných vrcholů, obklopených ledovcem). Jsou to zrnka hornin, prach a živé i mrtvé organismy. Díky nižšímu albedu (podílu odraženého a dopadajícího záření), než má okolní led, absorbuje více tepla a protává do povrchu ledovce. Tvoří tak jamky s plochým dnem a svislými stěnami, které jsou do určité úrovně naplněny vodou. Jamky mohou dosahovat hloubky až několika decimetrů a plochy v řádech od milimetrů po metry čtvereční. Mohou představovat 2–5 % povrchu ablační zóny ledovce čili oblasti, kde během letní sezony roztaje sníh na povrchu ledu.
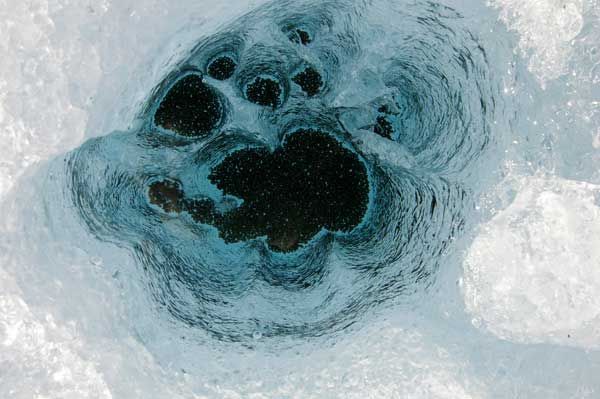 Povrch ledovce tu nabízí podmínky zcela neočekávané: Během letního období je tu stabilní teplota – i když prakticky jen na bodu tání okolního ledu – a neustále přítomná tekutá voda. Pevný substrát, na kterém se mají buňky „kde chytit“, umožňuje koncentraci živin a zároveň vytváří variabilitu světelných podmínek. V kryokonitu dominují prokaryotní organismy, především heterotrofní bakterie a sinice, vyskytuje se v něm ale i řada jednoduchých, zpravidla jednobuněčných zelených řas, rozsivek, kvasinek a prvoků ,a dokonce aktivní mnohobuněční živočichové jako vířníci (Rotifera), želvušky (Tardigrada) a chvostoskoci (Collembola).
Povrch ledovce tu nabízí podmínky zcela neočekávané: Během letního období je tu stabilní teplota – i když prakticky jen na bodu tání okolního ledu – a neustále přítomná tekutá voda. Pevný substrát, na kterém se mají buňky „kde chytit“, umožňuje koncentraci živin a zároveň vytváří variabilitu světelných podmínek. V kryokonitu dominují prokaryotní organismy, především heterotrofní bakterie a sinice, vyskytuje se v něm ale i řada jednoduchých, zpravidla jednobuněčných zelených řas, rozsivek, kvasinek a prvoků ,a dokonce aktivní mnohobuněční živočichové jako vířníci (Rotifera), želvušky (Tardigrada) a chvostoskoci (Collembola).
 Podobně jako je v kryokonitu pestrý život, jsou pestré i otázky, kterými zaměstnává vědeckou komunitu. V centru současného výzkumu leží metabolická aktivita tohoto prostředí, její možnosti pro přeměnu různých substrátů a možný dopad na globální bilanci uhlíku. Na základě současných poznatků víme, že metabolická aktivita na povrchu ledovců je dost velká na to, aby měl kryokonit při své rozloze význam v globálním měřítku. Vede se však stále diskuse o tom, zda je kryokonit v celkové bilanci heterotrofní nebo autotrofní systém, tedy zda se z povrchu ledovců uvolňuje CO2, nebo je uhlík naopak dlouhodobě ukládán ve formě organické hmoty. K dispozici je poměrně málo měření, která zároveň pokrývají jen období, kdy je kryokonit rozmrzlý. Spíš zatím tušíme, než víme, s jakou intenzitou může heterotrofická aktivita pokračovat na konci sezony, kdy kryokonit uzavře ledová poklička a ocitne se pod sněhem ve tmě.
Podobně jako je v kryokonitu pestrý život, jsou pestré i otázky, kterými zaměstnává vědeckou komunitu. V centru současného výzkumu leží metabolická aktivita tohoto prostředí, její možnosti pro přeměnu různých substrátů a možný dopad na globální bilanci uhlíku. Na základě současných poznatků víme, že metabolická aktivita na povrchu ledovců je dost velká na to, aby měl kryokonit při své rozloze význam v globálním měřítku. Vede se však stále diskuse o tom, zda je kryokonit v celkové bilanci heterotrofní nebo autotrofní systém, tedy zda se z povrchu ledovců uvolňuje CO2, nebo je uhlík naopak dlouhodobě ukládán ve formě organické hmoty. K dispozici je poměrně málo měření, která zároveň pokrývají jen období, kdy je kryokonit rozmrzlý. Spíš zatím tušíme, než víme, s jakou intenzitou může heterotrofická aktivita pokračovat na konci sezony, kdy kryokonit uzavře ledová poklička a ocitne se pod sněhem ve tmě.
Nyní opustíme tenkou vrstvu ledovce, kde je fotosyntéza alespoň sezonně možná, a sestoupíme do vrstev hluboko pod povrchem, kde jasně převládají chemotrofní typy metabolismu.
Englaciální biotop
Nitro ledovců patří mezi skutečně překvapivá místa. Životní pochody mohou probíhat pouze při dostupnosti tekuté vody, zatímco teploty v nitru ledovců klesají přibližně až na –30 °C. Buňky uvězněné v ledu jsou během procesu mrznutí a tvorby ledu vytěsňovány na křižovatky úzkých kanálků mezi jednotlivými zrny či krystaly (viz rovněž Vesmír 79, 253, 2000/5 a 79, 323, 2000/6). Tyto kanálky obsahují tekutou vodu i při velmi nízkých teplotách. Mohou za to ionty, které jsou v procesu tvorby krystalické mřížky ledu rovněž vytěsněny a jejichž koncentrace ve zbývající vodě roste s klesající teplotou ledu. Tekutá voda je také přítomná ve formě tenkého filmu na styku ledu a povrchu minerálů i v extrémně nízkých teplotách.
 Buňky (většinou pouze velmi malé prokaryotické) uvězněné v ledu mohou díky tomu vyměňovat s okolím látky a energii a zachovávat si tak životaschopnost. Intenzita metabolismu těchto ledových vězňů je natolik nízká, že i při velkém objemu ledu (ve srovnání s víceméně dvourozměrnými supraglaciálními a subglaciálními biotopy) nemají významný vliv na přeměnu látek a energie v ledovcových ekosystémech. Přesto mohou způsobovat pomalé změny koncentrací substrátů a produktů v malém lokálním měřítku – a to je třeba mít na paměti při analýzách velmi starých ledovcových jader.
Buňky (většinou pouze velmi malé prokaryotické) uvězněné v ledu mohou díky tomu vyměňovat s okolím látky a energii a zachovávat si tak životaschopnost. Intenzita metabolismu těchto ledových vězňů je natolik nízká, že i při velkém objemu ledu (ve srovnání s víceméně dvourozměrnými supraglaciálními a subglaciálními biotopy) nemají významný vliv na přeměnu látek a energie v ledovcových ekosystémech. Přesto mohou způsobovat pomalé změny koncentrací substrátů a produktů v malém lokálním měřítku – a to je třeba mít na paměti při analýzách velmi starých ledovcových jader.
V akumulačních zónách ledovců, kde díky morfologii ledovce nedocházelo k velkým posunům (například v centrálních oblastech velkých polárních ledovců), lze na vertikálním profilu rozlišit roční vrstvy akumulovaného sněhu přeměněného tlakem v led. Tato posloupnost vrstev představuje chronologický záznam srážek, pevných spadů i vzduchu uvězněného mezi krystaly stovky tisíc až miliony let a používá se při rekonstrukci změn podnebí. V ledových jádrech grónského ledovce (GISP2) byly nalezeny zvýšené koncentrace metanu, které jsou s největší pravděpodobností výsledkem činnosti metanogenních archebakterií při teplotě ledu až –9 °C.
Aktivní englaciální mikroorganismy nám tak přinášejí dvě poselství: Život neustává při 0 °C, ale může pokračovat při velmi nízkých teplotách, pokud je přítomná tekutá voda. A minulost naší planety zapsaná v hlubokých ledovcových profilech proto může být pomalu přepisována.
Subglaciální biotopy
Místa na kontaktu podloží a ledovcového dna mají také svá výrazná specifika. I když ledovce o malé tloušťce mají své dno typicky přimrzlé k podkladu, při určité mocnosti vrstvy ledu (kolem 100 m) tlak dostatečně sníží bod tání ledu a změní jeho konzistenci na rozhraní ledové báze s podložím. To zaprvé významně ovlivňuje dynamiku ledové masy (ledovec může po takové vrstvě klouzat) a zároveň poskytuje životní prostor mikroorganismům, kterým nevadí zima, tma, velký tlak a hladovění. Díky omezené výměně plynů s atmosférou tu mikroorganismy často vydýchají kyslík a vzniknou anoxické podmínky. Ke slovu se dostávají mikrobi s různými typy anaerobního metabolismu (denitrifikací, redukcí síranů a trojmocného železa, acetogenezí a metanogenezí).
 Subglaciální metanogeneze, tedy biogenní produkce nejúčinnějšího skleníkového plynu metanu pod ledovci je jedním z důležitých témat současného polárního výzkumu. Metanogenní archea – producenti metanu, jejichž nejbližší příbuzní žijí např. v trávicích traktech přežvýkavců či v anoxických mokřadech – byla nalezena v subglaciálních sedimentech na dně ledovců v Antarktidě, Grónsku či na Špicberkách a produkce metanu v těchto sedimentech byla potvrzena v laboratoři. Přestože intenzita procesu je v podledovcových podmínkách velmi nízká, vzhledem k času, který měli mikrobi k dispozici (antarktický ledový štít je starý více než 30 milionů let), může být množství metanu vyprodukovaného těmito společenstvy nezanedbatelné. Při nárazovém uvolnění do atmosféry mohl metan v minulosti způsobovat značné klimatické změny.
Subglaciální metanogeneze, tedy biogenní produkce nejúčinnějšího skleníkového plynu metanu pod ledovci je jedním z důležitých témat současného polárního výzkumu. Metanogenní archea – producenti metanu, jejichž nejbližší příbuzní žijí např. v trávicích traktech přežvýkavců či v anoxických mokřadech – byla nalezena v subglaciálních sedimentech na dně ledovců v Antarktidě, Grónsku či na Špicberkách a produkce metanu v těchto sedimentech byla potvrzena v laboratoři. Přestože intenzita procesu je v podledovcových podmínkách velmi nízká, vzhledem k času, který měli mikrobi k dispozici (antarktický ledový štít je starý více než 30 milionů let), může být množství metanu vyprodukovaného těmito společenstvy nezanedbatelné. Při nárazovém uvolnění do atmosféry mohl metan v minulosti způsobovat značné klimatické změny.
Permafrost
Trvale zmrzlá půda není součástí glaciálních systémů a najdeme ji i tam, kde je nedostatek srážek pro vznik ledovce (např. v rozsáhlých oblastech Sibiře či arktické Severní Ameriky). S ledovcovými biotopy má ale mnoho společného.
 Mikroorganismy (zejména prokaryota) uvězněné v permafrostu si mohou uchovat schopnost obnovy metabolismu a růstu po rozmrznutí i po miliony let (zatím nebyl objeven sterilní permafrost). V permafrostu je rovněž na rozsáhlých územích severní polokoule typický vysoký obsah nerozložené organické hmoty. Během tání permafrostu se rozbíhají biologické procesy a uvolňuje se velkého množství skleníkových plynů (CO2, CH4), což je notoricky známá pozitivní zpětná vazba posilující skleníkový efekt. Zároveň se do ekosystému mohou uvolňovat mikroorganismy (včetněvirů) uvězněné v permafrostu po dlouhá období. To může mít velký význam pro přenos viru na recentního hostitele nebo pro začlenění starého mikrobiálního genu do živé bakterie. Zatím o frekvenci takových přenosů příliš nevíme, protože jsou pro naše pozorovatelské možnosti vzácné. To ale neznamená, že se nedějí a nemůžou se stát ekologicky nebo evolučně významnou událostí.
Mikroorganismy (zejména prokaryota) uvězněné v permafrostu si mohou uchovat schopnost obnovy metabolismu a růstu po rozmrznutí i po miliony let (zatím nebyl objeven sterilní permafrost). V permafrostu je rovněž na rozsáhlých územích severní polokoule typický vysoký obsah nerozložené organické hmoty. Během tání permafrostu se rozbíhají biologické procesy a uvolňuje se velkého množství skleníkových plynů (CO2, CH4), což je notoricky známá pozitivní zpětná vazba posilující skleníkový efekt. Zároveň se do ekosystému mohou uvolňovat mikroorganismy (včetněvirů) uvězněné v permafrostu po dlouhá období. To může mít velký význam pro přenos viru na recentního hostitele nebo pro začlenění starého mikrobiálního genu do živé bakterie. Zatím o frekvenci takových přenosů příliš nevíme, protože jsou pro naše pozorovatelské možnosti vzácné. To ale neznamená, že se nedějí a nemůžou se stát ekologicky nebo evolučně významnou událostí.
Na povrchu aktivní vrstvy permafrostu se rozvíjejí půdní krusty tvořené sinicemi, řasami, houbami, lišejníky a mechy, samozřejmě spolu s bakteriemi a prvoky. Krusty představují první mechanickou stabilizaci substrátu a základ pozdějšího vývoje půdy. Jsou odolné k vysychání, ke kterému snadno dochází v nestabilních hydrologických poměrech na rozmrzlém povrchu permafrostu. Vyschnou- li, jejich úlomky snadno unáší vítr a stávají se pravděpodobně významným zdrojem mikrobiálních inokul a organické hmoty, přinášené na povrch ledovců.
Dry Valleys: Trvale zamrzlá jezera
Velmi specifické biotopy, kromě výše uvedených, lze nalézt v Antarktidě. Jsou jimi například trvale zamrzlá jezera v Suchých údolích (Dry Valleys), největším odledněném území antarktického kontinentu. Tato jezera se dramaticky liší od jakýchkoli jiných jezer naší planety celoroční přítomností několikametrové (okolo 3 až 8 m) ledové pokrývky. Pod touto vrstvou se ve studených vodách jezer ukrývá nevídaně bohatý ekosystém obývaný bakteriemi, sinicemi, řasami a drobnými vířníky.
Led na povrchu brání větru v míchání vody a v jednom jezeře proto na sobě mohou být navrstvené vody s rozdílnou salinitou, teplotou, chemickým složením i složením mikroorganismů. Zatímco hlubinné vrstvy jezer, kam už nedopadá světlo, bývají velice slané a mohou v nich panovat anaerobní podmínky, v horních vrstvách je voda sladká a daří se zde fytoplanktonu složenému především ze zelených řas.
Většina živin v těchto jezerech pochází ještě z doby před mnoha desítkami tisíc let, kdy Suchá údolí byla součástí Rossova moře. Novodobý přísun živin je minimální. Pouze v létě rozmrzají okraje ledu a jezera jsou napájena vodou z tajících ledovců. A někdy do nich také protaje navátý sediment podobně jako při vzniku kryokonitu.
Ledová pokrývka také do jisté míry chrání jezera před okolními mrazivými teplotami. Jezera v Suchých údolích jsou proto jediným známým biotopem Antarktidy, kde se kapalná voda vyskytuje během celého roku, tj. i v období polární zimy, kdy teploty klesají až k –65 °C. Co se děje s mikrorganismy v jezerech během dlouhých zimních měsíců, kdy se zde nenachází žádné světlo, není úplně jasné. Nejnovější výzkumy naznačují, že řada druhů fytoplanktonu, například kryptomonády (skrytěnky), se s ubývajícím množstvím světla přeorientovává z fotosyntézy na aktivní lov a konzumaci bakterií. To dokazuje, že organismy během zimy v Antarktidě nejen přežívají, ale dokonce se aktivně živí a rozmnožují. Život v Antarktidě se ani v zimě nezastavuje, jak se dříve předpokládalo.
Od doby, kdy Pyne napsal slova uvedená v úvodu, se naše poznání života v chladných podmínkách radikálně změnilo. Vděčíme za to především překotnému rozvoji molekulární biologie, která nejen změnila úhel pohledu, pod kterým je možné organismus zkoumat, ale umožnila řadu organismů (zejména prokaryotních) vůbec registrovat. Slovo registrovat nejlépe vystihuje povahu tohoto druhu poznání. Rychle přibývá nově objevených sekvencí DNA a dramaticky „bobtnají“ metagenomické databáze.
O „skutečných“ mikroorganismech však ve většině případů nic nevíme – jen to, že existují. Neumíme je zatím kultivovat a jen podle nejbližších nalezených příbuzných můžeme usuzovat, o jaké organismy jde. Dnes už nejen víme, že ledovce i kontinentální ledové příkrovy hostí živé organismy, ale jsme schopni měřit jejich metabolickou aktivitu a dokonce je identifikovat a kvantifikovat. Například pomocí fluorescenčně značených oligonukleotidů, které se vážou na specifické sekvence rRNA. Podle přítomnosti určitých genů rovněž můžeme odhadovat jejich schopnost ovlivňovat prostředí, v němž žijí.
O životě mikrobů ve volné přírodě, a nejen těch z chladných koutů biosféry, víme v porovnání s rostlinami nebo živočichy velice málo. Přitom pro tok látek a energie v přírodě je leckterá bakteriální populace významnější než mnohý druh obratlovce. Dalo by se říci, že náš „mnohobuněčný“ svět dosahuje při velmi uniformní kombinaci biochemických pochodů obrovské variability forem, zatímco mikrobiální svět si při minimální diverzitě tvarů udržuje dosud ne zcela probádané bohatství biochemických pochodů. Tyto pochody mají velký kvantitativní podíl především na využívání různých substrátů a oživení rozmanitých prostředí.
Adaptace na život s ledem
V případě jednobuněčnosti lze těžko mluvit o typické adaptaci, nicméně většina složitějších mnohobuněčných organismů zmrznutí a rozmrznutí nepřežije a jednoduchost jednobuněčného života představuje v chladu a mrazu velkou výhodu. Existují bezesporu podivuhodné výjimky například mezi hmyzem (střevlíček Pterostichus brevicornis, který je schopen přežít teplotu až –87 °C; viz Vesmír 88, 304, 2009/5, 88, 368, 2009/6 a 88, 480, 2009/7), ale v biotopech vázaných na led jasně dominuje mikrobiální život.
Život v nízkých teplotách vyvíjí tlak na řadu vlastností buněk. Zde vyjmenujeme ty základní, seřazené zhruba od krátkodobých „ekologických“ po dlouhodobé „evoluční“:
- Složení buněčných membrán je odlišné. Zvyšuje se podíl nenasycených mastných kyselin s kratšími řetězci. Díky tomu je i při nízkých teplotách zachována dostatečná fluidita membrán.
- V pravidelně zamrzajících a rozmrzajících společenstvech mají organismy tendenci k zmenšování rozměrů buněk nebo k syntéze a hromadění kryoprotektantů. Malá buňka znamená větší povrch vůči objemu, což umožní rychlejší ztrátu vody během koncentrace látek v okolním prostředí v důsledku tvorby vodních krystalů. Kryoprotektanty mohou být buď specifické a vázat se přímo na povrch vznikajících krystalů vně i uvnitř buněk a omezovat jejich rozměry, nebo jde jednoduše jen o malou molekulu (alkohol, aminokyselina) zvyšující osmotický potenciál buňky, což sníží bod tání protoplastu.
- Zvyšuje se flexibilita enzymů, což pro specialisty na této úrovní představuje vážný problém při vyšší teplotě.
- Oproti termofilním organismům je v genomu tendence k vyššímu podílu nukleotidových párů adenin–tymin (AT) oproti cytosin–guanin (GC ). To snižuje energii vazby mezi oběma řetězci DNA a vede ke snížení energetických nároků na replikaci.
Ke stažení
 článek ve formátu pdf [623,88 kB]
článek ve formátu pdf [623,88 kB]
O autorech
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
