









Kryptobiotické stavy aneb Vratná zastavení životních dějů
 Život je v našem chápání většinou spojen s dějem. Téměř každá definice života odlišuje živé objekty od neživých na základě přítomnosti aktivních životních procesů, jako jsou výměna energie, metabolismus, růst, vývoj a rozmnožování. Smrt potom chápeme jako nevratné zastavení životních procesů. V přírodě a v laboratoři se ovšem setkáváme také s organismy, u nichž je zastavení životních dějů vratné; jejich život se dočasně redukuje na pouhou životaschopnou strukturu. Jeden ze základních atributů života tedy není přítomen, ale o smrti také nelze mluvit, jelikož návrat k životu je možný. Mluvíme o anabióze („návratu do života“) nebo synonymicky o kryptobióze („skrytém životě“). Definice kryptobiotického stavu vyžaduje, aby jakékoli životní procesy organismu byly zcela zastaveny (teoreticky) nebo přinejmenším zpomaleny na neměřitelné hodnoty (prakticky). 1) Tím se kryptobióza liší od různých letargických a dormantních stavů, za kterých životní procesy nadále pokračují, byť ve zpomalené a různě modifikované podobě.
Život je v našem chápání většinou spojen s dějem. Téměř každá definice života odlišuje živé objekty od neživých na základě přítomnosti aktivních životních procesů, jako jsou výměna energie, metabolismus, růst, vývoj a rozmnožování. Smrt potom chápeme jako nevratné zastavení životních procesů. V přírodě a v laboratoři se ovšem setkáváme také s organismy, u nichž je zastavení životních dějů vratné; jejich život se dočasně redukuje na pouhou životaschopnou strukturu. Jeden ze základních atributů života tedy není přítomen, ale o smrti také nelze mluvit, jelikož návrat k životu je možný. Mluvíme o anabióze („návratu do života“) nebo synonymicky o kryptobióze („skrytém životě“). Definice kryptobiotického stavu vyžaduje, aby jakékoli životní procesy organismu byly zcela zastaveny (teoreticky) nebo přinejmenším zpomaleny na neměřitelné hodnoty (prakticky). 1) Tím se kryptobióza liší od různých letargických a dormantních stavů, za kterých životní procesy nadále pokračují, byť ve zpomalené a různě modifikované podobě.
Kryptobióza nabízí nejen úžasné téma pro obecné úvahy nad hranicemi mezi životem a smrtí či nad možností cestování prostorem a časem, ale také široký prostor pro základní výzkum mechanismů regulovaného pozastavení životních procesů. A základní výzkum dnes pomalu přechází do období prvních praktických aplikací v technologiích dlouhodobého skladování biologického materiálu nebo celých organismů.
V přírodě existují tři základní příčiny vstupu organismu do kryptobiotického stavu: nedostatek vody (anhydrobióza), nízká teplota (kryobióza) a nedostatek energie (anoxybióza). Postupně představíme principy všech tří stavů, přičemž se soustředíme zejména na situaci u živočichů (Metazoa).
Anhydrobióza Nejmarkantnějším příkladem kryptobiózy je patrně anhydrobióza – stav způsobený nedostatkem vody v organismu (viz rámeček Trocha suché historie 1 ).
Vodné prostředí umožňuje pohyb molekul.
 Mohou se přesunovat, přenášet hmotu, energii a informaci z místa na místo a vstupovat do vzájemných interakcí. Klesne-li podíl vody v organismu pod určitou kritickou mez (uvádějí se hodnoty 0,1–0,3 g vody na gram sušiny), je možnost pohybu molekulám téměř odepřena. Právě tuto mez můžeme považovat za hranici anhydrobiózy. Není třeba, aby obsah vody klesl až k nule; bez možnosti pohybu molekul se zastaví většina biochemických procesů a s nimi i (měřitelný) život.
Mohou se přesunovat, přenášet hmotu, energii a informaci z místa na místo a vstupovat do vzájemných interakcí. Klesne-li podíl vody v organismu pod určitou kritickou mez (uvádějí se hodnoty 0,1–0,3 g vody na gram sušiny), je možnost pohybu molekulám téměř odepřena. Právě tuto mez můžeme považovat za hranici anhydrobiózy. Není třeba, aby obsah vody klesl až k nule; bez možnosti pohybu molekul se zastaví většina biochemických procesů a s nimi i (měřitelný) život.
Schopnost přežít v anhydrobiotickém stavu rozhodně není univerzální. Naopak složitější organismy jako obratlovci nebo vyšší rostliny jsou velmi citlivé i na mírný pokles obsahu vody v těle. To platí i pro učebnicové mistry v přežívání sucha, pro africké a jihoamerické bahníky nebo australské pouštní žáby. Přežijí sice několikaměsíční vymizení vody z jejich prostředí, sami ovšem nevysychají. Pouze vstupují do stavu hluboké dormance, během níž vystačí se zásobou vody v těle. Schopnost přežít v anhydrobiotickém stavu je vlastní pouze vybraným organismům, a pokud bychom hledali jejich společného jmenovatele, tak organismům malým. Malá velikost těla jednoznačně limituje možnost vytvořit si dostatečnou zásobu vody, která při reálné propustnosti tělní slupky a nepříznivém poměru objemu vůči povrchu vystačí na rozumnou dobu (jež odpovídá evoluční zkušenosti s periodickým suchem).
 Anhydrobiontům je rovněž společná jednoduchost. Přežije-li vyschnutí jednotlivá buňka nebo embryo, příliš nás to nepřekvapí. Ostatně použití suchých kvasinek v kuchyni je běžné a lidská společnost závisí na skladování rostlinných semen v suchém stavu. Výčet anhydrobiontů ale zahrnuje kromě jednoduchých bakterií, kvasinek, řas, lišejníků, mechorostů, rostlinných semen a embryonálních cyst různých bezobratlých také poměrně složité organismy, jako jsou některé cévnaté rostliny, hlístice, vířníci, želvušky, a dokonce hmyz. Larvy pakomára Polypedilum vanderplancki 2) jsou největším a nejsložitějším známým organismem, který je schopen přežít téměř úplnou ztrátu vody z těla. Mají plně vyvinutou nervovou soustavu, smysly, svaly, žlázy, trávicí trakt, vylučovací ústrojí ap. Pokud je tedy složitost organismu vůbec nějakým omezením pro anhydrobiózu, pak je její limit stanoven překvapivě vysoko.
Anhydrobiontům je rovněž společná jednoduchost. Přežije-li vyschnutí jednotlivá buňka nebo embryo, příliš nás to nepřekvapí. Ostatně použití suchých kvasinek v kuchyni je běžné a lidská společnost závisí na skladování rostlinných semen v suchém stavu. Výčet anhydrobiontů ale zahrnuje kromě jednoduchých bakterií, kvasinek, řas, lišejníků, mechorostů, rostlinných semen a embryonálních cyst různých bezobratlých také poměrně složité organismy, jako jsou některé cévnaté rostliny, hlístice, vířníci, želvušky, a dokonce hmyz. Larvy pakomára Polypedilum vanderplancki 2) jsou největším a nejsložitějším známým organismem, který je schopen přežít téměř úplnou ztrátu vody z těla. Mají plně vyvinutou nervovou soustavu, smysly, svaly, žlázy, trávicí trakt, vylučovací ústrojí ap. Pokud je tedy složitost organismu vůbec nějakým omezením pro anhydrobiózu, pak je její limit stanoven překvapivě vysoko.
Případ sušených pakomářích larev
Pakomáři P. vanderplancki obývají africký sahel. Samičky kladou oplodněná vajíčka do stojaté vody různých tůněk a louží. Mimo jiné také do dočasných jezírek, která se vytvářejí v období dešťů ve skalních prohlubních. Larvy procházejí obvyklým procesem růstu a larválních svlékání až do stadia kukly, ze které se opět líhnou dospělí pakomáři. Celý vývojový cyklus se dá stihnout během měsíce nebo dvou. Tedy za předpokladu, že voda nevyschne. To se ale právě malým skalním loužím stává poměrně často. Larvy komárů rodu Chironomus, které někdy obývají louži spolu s larvami pakomárů, vyschnou a nepřežijí (viz rámeček Kanibalistické východisko z problému 2 ).
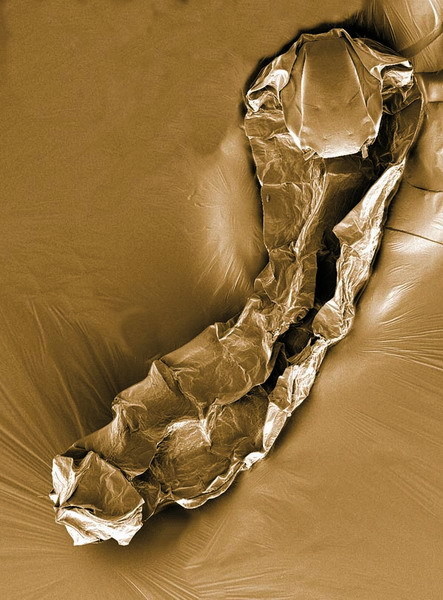
Larvy pakomárů P. vanderplancki také vyschnou a promění se v útvar, který vzhledem i konzistencí připomíná zrnko písku (obrázek 1). A při měření jakýchkoli životních projevů se jako zrnko písku rovněž chová; nenaměříme nic ani pomocí nejcitlivějších přístrojů. Na první pohled tedy larvy komárů a pakomárů skončí stejně špatně. Zásadní rozdíl se však projeví v příštím období dešťů. Larvy pakomárů během několika desítek minut nasáknou vodou a opět ožijí! Celý proces vyschnutí a následného zavodnění můžeme snadno simulovat v laboratoři (obrázek 2). Larvy umístíme do Petriho misky s malým množstvím vody a vodu necháme pomalu odpařovat (pomalé tempo odpařování dává larvám šanci zahájit a dokončit fyziologickou přípravu na vyschnutí). Výsledkem bude objekt s relativním obsahem vody kolem 2 %, hluboko pod mezní hranicí pro aktivní biochemické procesy. Pokud jsou anhydrobiotické larvy skladovány v suchu, temnu a při pokojové teplotě, vydrží minimálně 19 let. Tak dlouhý je dosud nejdelší sledovaný záznam, který ovšem dál pokračuje. 3) Maximální doba skladování bude zřejmě mnohonásobně delší.
 Anhydrobiotické organismy vydrží ovšem i leccos jiného: zchlazení na teplotu kapalného helia (–270 °C), zahřátí na teplotu 150 °C, koupel v 100% alkoholu nebo v organických rozpouštědlech, pobyt ve vakuu, vysoké dávky rentgenového záření či tlak 6000 atmosfér. Tyto extrémní výkony přivedly některé vědce k myšlence testovat schopnost přežití anhydrobiotů v otevřeném vesmíru. První úspěšná mise proběhla v září 2007, kdy raketa Evropské vesmírné agentury vynesla na oběžnou dráhu dva druhy želvušek (Tardigrada) v anhydrobiotickém stavu. Želvušky byly na 10 dnů vystaveny působení vakua, UV radiace a kosmického záření. 4) Přežily! Další test v kosmu právě probíhá, a to právě s anhydrobitickými pakomáry. Jejich plánovaný měsíční pobyt v otevřeném prostoru se kvůli hurikánu nakonec protáhl na celý rok. Američtí astronauti museli opustit oběžnou dráhu o den dříve a nestihli již odmontovat z vnějšího pláště speciální kontejner s pakomáronauty. Podle ústního sdělení vedoucího výzkumu Takashiho Okudy (Tsukuba, Japonsko) přesto někteří pakomáři přežili.
Anhydrobiotické organismy vydrží ovšem i leccos jiného: zchlazení na teplotu kapalného helia (–270 °C), zahřátí na teplotu 150 °C, koupel v 100% alkoholu nebo v organických rozpouštědlech, pobyt ve vakuu, vysoké dávky rentgenového záření či tlak 6000 atmosfér. Tyto extrémní výkony přivedly některé vědce k myšlence testovat schopnost přežití anhydrobiotů v otevřeném vesmíru. První úspěšná mise proběhla v září 2007, kdy raketa Evropské vesmírné agentury vynesla na oběžnou dráhu dva druhy želvušek (Tardigrada) v anhydrobiotickém stavu. Želvušky byly na 10 dnů vystaveny působení vakua, UV radiace a kosmického záření. 4) Přežily! Další test v kosmu právě probíhá, a to právě s anhydrobitickými pakomáry. Jejich plánovaný měsíční pobyt v otevřeném prostoru se kvůli hurikánu nakonec protáhl na celý rok. Američtí astronauti museli opustit oběžnou dráhu o den dříve a nestihli již odmontovat z vnějšího pláště speciální kontejner s pakomáronauty. Podle ústního sdělení vedoucího výzkumu Takashiho Okudy (Tsukuba, Japonsko) přesto někteří pakomáři přežili.
Fyziologie anhydrobiózy
Zmíněné vlastnosti činí z anhydrobiotických organismů nesmírně zajímavé objekty základního výzkumu. Postupná dehydratace aktivuje biosyntézu a hromadění jednoduchých cukerných látek, jež jsou považovány za jeden z klíčů k úspěšnému přežití v anhydrobiotickém stavu. U rostlin je to především sacharóza, někdy doplněná menším množstvím rafinózy a stachyózy. Kvasinky a bezobratlí živočichové spoléhají na trehalózu. V anhydrobiotickém stavu může podíl trehalózy činit až 20 % celkové hmotnosti těla. U larev pakomárů je to kolem 18 %. Role trehalózy (a sacharidů obecně) není dosud plně objasněna. Patrně jde o kombinaci principů, z nichž některé již získaly dobrou teoretickou i empirickou podporu.
Pozitivní působení trehalózy začíná ještě před dosažením hranice anhydrobiotického stavu. Termodynamické vlastnosti systému voda-trehalóza-protein (membrána) znesnadňují denaturaci proteinů (popřípadě změnu vlastností membrány) při vzrůstající koncentraci destabilizujících iontů v roztoku, ze kterého postupně mizí voda. Molekuly trehalózy jsou přednostně vytěsňovány z bezprostřední blízkosti proteinů a biologických membrán, jejichž povrch naopak zůstává hydratovaný. 5) Pokračující dehydratace vede až ke ztrátám molekul vody přímo z hydratačních obalů proteinů a membrán. V prostředí bez trehalózy by to znamenalo fatální ztrátu funkčního prostorového uspořádání. Proteiny by denaturovaly a agregovaly, membrány by přešly do gelové nebo hexagonální fáze. Mezi trehalózou a povrchem proteinu či membrány se však vytvoří vodíkové můstky, jež nahradí chybějící molekuly vody. 6) Koncentrovaný roztok trehalózy je navíc silně viskózní a může přecházet až do amorfní sklovité struktury. 7) Biochemické procesy se zastavují, volný prostor po molekulách vody vyplňuje trehalóza. Delikátní struktura buněk a orgánů je chráněna před kolapsem a konzervována ve „ztuhlém“ stavu.
Ačkoli se současný základní výzkum dosud soustředil převážně na trehalózu, celá věc je složitější. Například někteří vířníci a některé želvušky úspěšně překonávají anhydrobiotickou epizodu, aniž k tomu potřebují trehalózu nebo jiný sacharid. Anhydrobióza se tak jeví jako složitý adaptivní jev, jehož je trehalóza pouhým jedním symptomem. Zdá se, že vlastnosti velmi podobné trehalóze mají i některé proteiny. LEA proteiny (Late Embryo Abundant) byly poprvé nalezeny v semenech rostlin, kde se ve velkém množství hromadí v pozdních fázích embryonálního vývoje, jež jsou spojeny s postupnou dehydratací. Později byly zaznamenány i v pylu a vegetativních orgánech rostlin a také v řasách a sinicích vystavených suchu. Nález LEA proteinů u vysychajících hlístic a vířníků potvrdil jejich rozšíření i mezi živočichy.
LEA proteiny jsou poměrně malé a silně hydrofilní. Mohou proto vázat ionty koncentrující se ve vysychající buňce, zvyšovat viskozitu prostředí a podporovat přechod do skelné fáze. 8) Interagují rovněž s povrchem proteinů a membrán a konzervují jejich strukturu. Kromě toho patrně (stejně jako trehalóza) vychytávají volné kyslíkové radikály. To je důležité, neboť ochrana proti oxidativnímu poškození je dalším významným opatřením pro přežití v dehydratovaném stavu. Tento ochranný systém a také systém šokových proteinů jsou obvykle aktivovány nejen při vysychání, ale i při různých jiných stresových stavech, do kterých se živé buňky dostávají.
Praktické aplikace na obzoru
Využití trehalózy při dlouhodobém skladování buněk, orgánů nebo celých organismů v suchém stavu a za pokojových teplot zní jako velmi lákavá alternativa k nákladnému a složitému uchovávání hluboko pod bodem mrazu. Současný aplikovaný výzkum již dosáhl povzbudivých, i když omezených úspěchů při skladování savčích červených krvinek, lidských krevních destiček, kmenových buněk a spermatu. 9) Savčí buňky však neumějí vyrábět trehalózu. Navíc trehalóza jen velmi pomalu prochází biologickými membránami a při vnější aplikaci se do nitra buněk nedostane. K překonání těchto potíží používají vědci různé triky – přechodně zvyšují propustnost membrán během fázového přechodu při poklesu teploty, včleňují bakteriální geny pro biosyntézu trehalózy do genomu savčích buněk nebo pomocí zvláštního proteinu vytvářejí v membráně nespecifický pór. Jeden z nejnovějších triků odkoukali odborníci z laboratoře Takashiho Okudy od pakomářích larev.
V buňkách tukového tělesa dehydratovaných pakomářích larev se nejen rychle syntetizuje trehalóza, ale zároveň její specifický membránový přenašeč TRET1, který usnadňuje difuzi trehalózy ven z buněk. Okudův tým včlenil gen kódující TRET1 do genomu různých obratlovčích buněk (žabích oocytů, myších fibroblastů, křeččích ovariálních buněk či lidských jaterních buněk) a prokázal, že produkovaný protein v nich může fungovat jako účinný, specifický a obousměrný přenašeč trehalózy. 10) Možná jsme tak svědky přerůstání základního výzkumu pakomárů do praktických aplikací v lidské medicíně.
Poznámky
První pozorování anhydrobiotických organismů pochází od holandského přírodovědce Antoniho van Leeuwenhoeka. Roku 1702 sbíral sedimenty z okapů, pět měsíců je vysoušel, a potom znovu zalil převařenou vodou. Podle vlastních slov pak pod mikroskopem pozoroval „animalcula, […] která krátce po zalití začala rozpínat svá těla a po půl hodině aktivně plavat ve sklenici“. Dnes máme za to, že šlo o zástupce vířníků (Rotatoria). Pozorování anhydrobiotických hlístic učinili v polovině 18. století také John Needham a Henry Baker. Druhý z nich dokonce prohlásil: „Pozorujeme případ, kdy je život dočasně přerušen a zdánlivě zničen, […] aby po dlouhé době opět začal v témže těle […].“ Snad i tato vyjádření, která se přímo dotýkala otázek života, smrti a vzkříšení, podnítila širší zájem vědců.
Lazzaro Spallanzani, profesor pařížské univerzity a jeden z nejuznávanějších přírodovědců své doby, nejprve odmítl uznat pozorování anhydrobiotických hlístic za věrohodná. Po vlastních experimentech je ovšem nejen potvrdil, ale ještě rozšířil o záznamy vysoké odolnosti anhydrobiontů vůči vysoké a nízké teplotě. Různé body tehdejších diskusí ostatně přetrvávají dodnes. Především otázka úplného zastavení metabolismu stále vyvolává kontroverze.
Kanibalistické východisko z problému
Zajímavou alternativní strategii přežití v dočasně vyschlé africké louži používají larvy komárů rodu Chironomus. 12) Protože nedokážou přežít v anhydrobiotickém stavu, musí se spolehnout na urychlenou emigraci ještě před úplným vyschnutím dočasné vodní plochy. Nahuštění larev a chemické změny v odpařující se kaluži rozdělí populaci larev do dvou kohort. Kohorta „obrů“ pokračuje v urychleném vývoji. Dospělci dorůstají nadprůměrné velikosti, jež je vhodná pro přesun na nové vhodné stanoviště. Rychlý vývoj a růst obrů jde na úkor kohorty „trpaslíků“, kteří na další vývoj naopak rezignují, „dobrovolně“ hladovějí a umírají. Obři tím získají víc místa a zároveň další potravu z rozkládajících se těl. Sebeobětování trpaslíků lze pochopit jako altruistický čin na podporu genotypu sdíleného s obry, protože larvy obývající jednu louži často pocházejí od jediné samice.Ke stažení
 článek ve formátu pdf [292,14 kB]
článek ve formátu pdf [292,14 kB]
O autorech
František Weyda
Vladimír Košťál
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
