









Molekuly na povel III.
Díky pilné práci novinářů bulvárních i seriózních plátků má běžný občan vcelku vyhraněnou představu o molekulární genetice i o molekulárním biologovi. Každému se snadno vybaví slovní spojení jako kód života, tajemná DNA, klíčová molekula ap. Molekuly nukleových kyselin jsou nepochybně v přírodě velmi důležité a objevy s tím spojené zrychlí tep každému zasvěcenci. Nicméně podobně jako všednodenní život Maty Hari nemusel být nijak zábavný a ona sama mohla být nudná a znuděná, o molekulární biologii platí, že každodenní rutinní práce mnoho vzrušení nenabízí. V jedné situaci však zavládne v každé laboratoři hmatatelné napětí – když se snažíme prokázat, že jsme skutečně dospěli ke kýženému výsledku.
 Ona totiž DNA, ačkoliv je molekulou vpravdě obrovskou, není běžně viditelná. Pokud s ní tedy něco provádíme, musíme mít předem vymyšleno, jak si výsledek provádění ověřit. Vždycky je to metoda časově i přístrojově náročná a tak nějak hodně za roh. Jako byste přítomnost někoho u vás v bytě zjišťovali měřením otřesů předmětů na kuchyňském stole u souseda. I zde však platí, že základní princip není obtížné pochopit – no, a pak už si jenom vymýšlíme variace.
Ona totiž DNA, ačkoliv je molekulou vpravdě obrovskou, není běžně viditelná. Pokud s ní tedy něco provádíme, musíme mít předem vymyšleno, jak si výsledek provádění ověřit. Vždycky je to metoda časově i přístrojově náročná a tak nějak hodně za roh. Jako byste přítomnost někoho u vás v bytě zjišťovali měřením otřesů předmětů na kuchyňském stole u souseda. I zde však platí, že základní princip není obtížné pochopit – no, a pak už si jenom vymýšlíme variace.
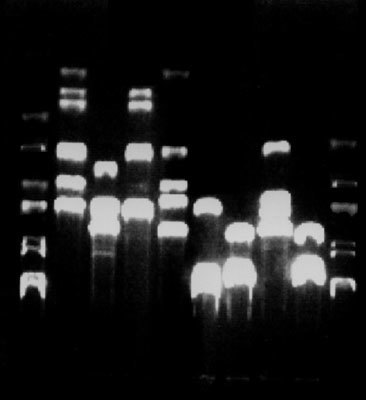 Zviditelněním svého druhu je vyhledávání pomocí sondy při hybridizaci (viz Vesmír 77, 312, 1998/6). Pokud získáme na filmu černý flek, naše DNA je ve vzorku – vidíme ji tam. Zcela zvláštní typ „vidění“ je prostřednictvím bakterií. DNA vneseme do bakterií a takto upravené buňky vysejeme na médium, kde mohou vyrůst jenom buňky s cizorodou DNA v bříšku. Pokud tedy bakterie vyrostou, je v nich ta správná DNA a díky bakteriím ji vidíme. Odtud pramení ten poněkud přezíravý vztah molekulárních biologů k bakterii Escherichia coli, která k takovým účelům slouží. Nejčastějším způsobem, jak si DNA prohlédnout, však zůstává elektroforéza.
Zviditelněním svého druhu je vyhledávání pomocí sondy při hybridizaci (viz Vesmír 77, 312, 1998/6). Pokud získáme na filmu černý flek, naše DNA je ve vzorku – vidíme ji tam. Zcela zvláštní typ „vidění“ je prostřednictvím bakterií. DNA vneseme do bakterií a takto upravené buňky vysejeme na médium, kde mohou vyrůst jenom buňky s cizorodou DNA v bříšku. Pokud tedy bakterie vyrostou, je v nich ta správná DNA a díky bakteriím ji vidíme. Odtud pramení ten poněkud přezíravý vztah molekulárních biologů k bakterii Escherichia coli, která k takovým účelům slouží. Nejčastějším způsobem, jak si DNA prohlédnout, však zůstává elektroforéza.
Hlavně elektřina
Molekulární biologie, tak jako kterákoli jiná činnost člověka, jede hlavně na elektřinu. Nukleové kyseliny jsou totiž skutečně kyseliny, tedy záporně nabité molekuly. Za jejich (nemalý) záporný náboj mohou fosfátové spojky mezi jednotlivými nukleotidy. Díky tomu, že jsou negativně nabité, pohybují se v elektrickém poli stejnosměrného proudu vždy od katody k anodě, tedy od – k +. To samo o sobě ale ještě nestačí. Molekuly nukleových kyselin se nepohybují jen tak ve vodě nebo v nějakém vodivém roztoku. Obvykle je necháme prolézat gelem, který se připravuje například z agarózy nebo z polyakrylamidu.
 Elektroforézou molekuly sice rozdělíme, máme je na gelu, ale stejně nic nevidíme. Proto je nutné gel s našimi vzorky obarvit. To se dělá pomocí vmezeřující barvičky etidiumbromidu. Tato barvička se skutečně vmezeří do záhybů dvoušroubovice DNA a zůstane tam poměrně pevně zaklíněna – a pořád ještě nic nevidíme. Teprve když na takto obarvenou DNA posvítíme ultrafialovým světlem, vmezeřená barvička začne krásně oranžově svítit. A svítí jenom tam, kde nějaká DNA je (obr. obrázek).
Elektroforézou molekuly sice rozdělíme, máme je na gelu, ale stejně nic nevidíme. Proto je nutné gel s našimi vzorky obarvit. To se dělá pomocí vmezeřující barvičky etidiumbromidu. Tato barvička se skutečně vmezeří do záhybů dvoušroubovice DNA a zůstane tam poměrně pevně zaklíněna – a pořád ještě nic nevidíme. Teprve když na takto obarvenou DNA posvítíme ultrafialovým světlem, vmezeřená barvička začne krásně oranžově svítit. A svítí jenom tam, kde nějaká DNA je (obr. obrázek).
Variace na známá témata
Ani bych snad neměla psát, co všechno nám elektroforéza nukleových kyselin umožňuje, aby se vám z toho nezamotala hlava. Nicméně alespoň základní věci. Díky elektroforéze lze poměrně přesně určit velikost molekuly. Máte-li k dispozici molekulu (nebo radši více molekul) o známé velikosti, můžete s její pomocí určit velikost neznámé molekuly. Zhotovíte si kalibrační křivku (pro každý gel je jiná, neboť velice záleží na konkrétních podmínkách elektroforézy), kde osa x určuje velikost molekuly a osa y logaritmus překonané vzdálenosti. Z této kalibrační křivky pak odečítáte podle vzdálenosti od jamky i velikost neznámých molekul. Pokud máme nějakou větší molekulu a chceme se v ní alespoň trošku zorientovat, použijeme tzv. restrikční mapování. O restriktázách už víme, že nám mohou každou molekulu DNA rozštěpit na menší fragmenty – pokud v té molekule najdou svá příslušná restrikční místa. Naštěpíme-li si tedy molekulu DNA jednotlivými restriktázami i kombinacemi několika restriktáz, a pak naštěpenou DNA proženeme gelem při elektroforéze, získáme obrázek proužků DNA. Odečtením podle kalibrační křivky určíme jejich velikost, součtem velikostí všech fragmentů zjistíme celkovou velikost molekuly a vzájemným kombinováním štěpů jednotlivými restriktázami i kombinací restriktáz získáme celkový obraz o místech pro jednotlivé enzymy na molekule – vytvoříme restrikční mapu (obr. obrázek). Ta nám pak poskytuje základní orientaci pro danou molekulu. Elektroforéza nám od sebe oddělí jednotlivé fragmenty molekul nukleových kyselin. Tyto fragmenty můžeme z gelu izolovat a dále s nimi pracovat. Elektroforézou (ve zcela speciálním uspořádání) můžeme analyzovat i celé chromozomy. Také můžeme provést elektroforézu RNA. S tím si prostý výzkumník užije pořádný kus legrace, protože, jak už víme, RNA je třeba tvrdě bránit před RNázami.
Elektroforéza nám od sebe oddělí jednotlivé fragmenty molekul nukleových kyselin. Tyto fragmenty můžeme z gelu izolovat a dále s nimi pracovat. Elektroforézou (ve zcela speciálním uspořádání) můžeme analyzovat i celé chromozomy. Také můžeme provést elektroforézu RNA. S tím si prostý výzkumník užije pořádný kus legrace, protože, jak už víme, RNA je třeba tvrdě bránit před RNázami.
Elektroforeticky můžeme analyzovat i proteiny. Tam je to ovšem trochu složitější. Na rozdíl od nukleových kyselin nemají bílkoviny vyhraněný elektrický náboj, ale jejich celkový náboj záleží na aminokyselinovém složení. Některé buněčné bílkoviny jsou tedy pozitivně nabité, jiné negativně, některé více, jiné méně. Jejich pohyb gelem není určen pouze velikostí, ale i nábojem. Pro některé typy analýzy to vůbec nevadí, ale někdy to je velká nevýhoda. Lze ji ovšem odstranit tak, že elektroforézu proteinů provedeme v nadbytku látky sodiumdodecylsulfátu (SDS). To je taková celkem nezajímavá molekula, záporně nabitá, s dlouhým, dvanáctiuhlíkatým ocáskem. Jakmile potká nějakou bílkovinu, tak se na ni namotá. Tím na ni ovšem přenese i svůj záporný náboj, takže nám odpadne starost s rozdílným nábojem bílkovin a ty se při elektroforéze za těchto podmínek dělí jenom podle velikosti. Takto upravené bílkoviny pak proženeme polyakrylamidovým gelem, fixujeme, obarvíme – a už je vidíme!
Nejúžasnější věci však zjistíme kombinací všech nám dosud známých metod. Můžeme například naštěpit DNA různými restriktázami a vzorky analyzovat elektroforézou. DNA se rozdělí podle velikosti a my ji z gelu přeneseme na membránu pomocí kapilárních sil v procesu, jenž se česky (roztomile staromilecky) jmenuje přesávka, ale v laboratorní hantýrce mu nikdo neřekne jinak než blotování. DNA přenesenou na membráně (a během přenosu denaturovanou) pak hybridizujeme se sondou vyrobenou podle nějakého fragmentu, který v naší molekule musí být. Díky tomuto postupu získáme nejenom restrikční mapu, ale i přesnou lokalizaci nějakého pro nás zajímavého fragmentu v analyzované molekule (obr. obrázek). Nebo izolujeme veškerou buněčnou RNA, proženeme ji gelem, přeneseme na membránu a hybridizujeme se sondou vytvořenou podle sekvence nějakého úplně konkrétního, pro nás zajímavého genu. Získáme-li potom na filmu černý flek, znamená to, že v buňkách je přítomna RNA vytvořená podle onoho genu, a tedy nejenže je v genomu gen přítomen, ale navíc je i aktivní, tedy vytváří se podle něj mRNA (a zřejmě i bílkovina). Ale hlavně je elektroforéza opravdu jediná metoda, která umožňuje snadno a rychle vidět studovanou molekulu. A to není málo.
Jak se dělá elektroforéza?
Existuje samozřejmě několik možných způsobů. My si povíme jenom o jednom (všechny ostatní jsou do značné míry podobné). Do speciální formy na gely nalijeme roztok a vsuneme do ní tzv. hřeben. Po utuhnutí a vyndání hřebene získáme obdélník gelu s jamkami na nanesení vzorku (obr. Obrázek). Roztok DNA s modrou barvičkou (která je také záporně nabitá a ukazuje nám, zda vůbec a jak se záporně nabité molekuly gelem pohybují) naneseme do jamek, zapneme elektrický proud (stejnosměrný) a čekáme. Zatímco my neděláme nic, molekuly nukleových kyselin se proplétají vlákny, dutinkami, komůrkami a jeskyňkami gelu. Čím větší molekula, tím hůře jí to jde, a tak za určitou dobu překoná mnohem kratší vzdálenost než molekula malá, která se dutinkami gelu snadno prosmýkne. A nejen to. Například kruhová, pevně smotaná molekula se pohybuje gelem mnohem rychleji než stejně velká molekula lineární. Zkrátka na gelu se nám jednotlivé molekuly od sebe oddělí buď podle velikosti, nebo podle trojrozměrné struktury. A to je to, co nás zajímá.
Ke stažení
 Článek ve formátu PDF [196,65 kB]
Článek ve formátu PDF [196,65 kB]
O autorovi
Zuzana Storchová
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
