










Molekuly na povel IV.
V našem rychlokurzu metod molekulární biologie již pozorný čtenář pokročil tak daleko, že by se bez větších problémů mohl uplatnit v leckteré moderní laboratoři – přinejmenším jako teoretik a univerzální poradce. Dovede připravit elektroforézu a analyzovat na ní vzorky nukleových kyselin či bílkovin (Vesmír 77, 372, 1998/7). Dovede DNA štěpit a v případě potřeby i vhodně spojovat, což v konečném důsledku vede k vytvoření molekul rekombinantní DNA a například k produkci savčích bílkovin v bakteriích (Vesmír 77, 257, 1998/5 ). A konečně dovede sondou vyhledat obzvláště oblíbený úsek nukleové kyseliny v jakémkoliv genetickém materiálu či hybridizací DNA určit míru podobnosti DNA různých organizmů (Vesmír 77, 312, 1998/6).
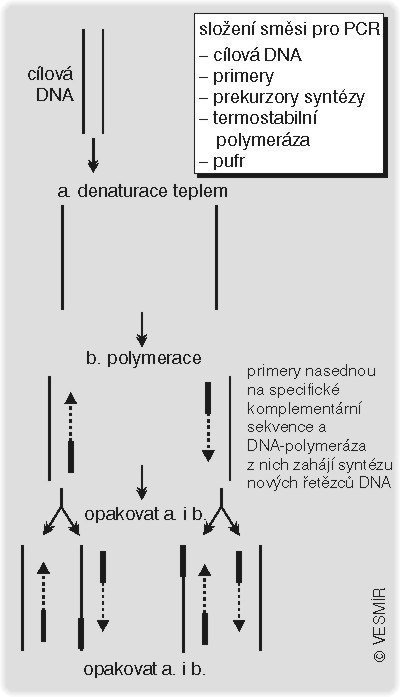 Všechny tyto metody jsou poměrně účinné, ale k jejich provedení je zapotřebí značné množství DNA. Abychom vzorek na elektroforéze dobře viděli, potřebujeme minimálně 5 (a spíše 50) ng DNA. Pomocí sondy dovedeme sice vyhledat i ještě menší množství DNA, ale čím menší a méně častý úsek DNA hledáte, tím větší musí být množství prohledávaného materiálu. A tak je velmi často nejdůležitější zajistit si pro práci dostatečné množství nukleových kyselin. Někdy to je docela snadné – stačí namnožit buňky a žádanou DNA z nich izolovat. To lze provést nejsnáze s bakteriemi, jež se pro pomnožení různých plazmidů (malých kruhových molekul DNA, do nichž můžeme vkládat úseky, které nás zajímají) používají nejčastěji. Bakterie v tomto případě nejsou terčem plánovaných pokusů, ale pouze jakýmsi laciným zdrojem požadovaného materiálu. Jestliže v nich chceme náš oblíbený fragment (NOF) pomnožit, potřebujeme jej vložit do plazmidu. Metody stříhání a opětovného slepování molekul DNA jsme sice probrali v jediné krátké lekci (viz Molekuly na povel I., Vesmír 77, 257, 1998/8), ale úplně snadné ani rychlé to není. Navíc ne vždy máte NOF připravený v pohotovosti pro vkládání do plazmidu – někdy ani nevíte, kde v chromozomu vlastně NOF leží. Proto objev postupu jak NOF namnožit do obrovského množství bez mezikroku v bakteriích pohnul výzkumem mohutně kupředu. Tato báječná technika se jmenuje polymerázová řetězová reakce (viz také Vesmír 69, 309, 1990/6; 74, 134, 1995/3).
Všechny tyto metody jsou poměrně účinné, ale k jejich provedení je zapotřebí značné množství DNA. Abychom vzorek na elektroforéze dobře viděli, potřebujeme minimálně 5 (a spíše 50) ng DNA. Pomocí sondy dovedeme sice vyhledat i ještě menší množství DNA, ale čím menší a méně častý úsek DNA hledáte, tím větší musí být množství prohledávaného materiálu. A tak je velmi často nejdůležitější zajistit si pro práci dostatečné množství nukleových kyselin. Někdy to je docela snadné – stačí namnožit buňky a žádanou DNA z nich izolovat. To lze provést nejsnáze s bakteriemi, jež se pro pomnožení různých plazmidů (malých kruhových molekul DNA, do nichž můžeme vkládat úseky, které nás zajímají) používají nejčastěji. Bakterie v tomto případě nejsou terčem plánovaných pokusů, ale pouze jakýmsi laciným zdrojem požadovaného materiálu. Jestliže v nich chceme náš oblíbený fragment (NOF) pomnožit, potřebujeme jej vložit do plazmidu. Metody stříhání a opětovného slepování molekul DNA jsme sice probrali v jediné krátké lekci (viz Molekuly na povel I., Vesmír 77, 257, 1998/8), ale úplně snadné ani rychlé to není. Navíc ne vždy máte NOF připravený v pohotovosti pro vkládání do plazmidu – někdy ani nevíte, kde v chromozomu vlastně NOF leží. Proto objev postupu jak NOF namnožit do obrovského množství bez mezikroku v bakteriích pohnul výzkumem mohutně kupředu. Tato báječná technika se jmenuje polymerázová řetězová reakce (viz také Vesmír 69, 309, 1990/6; 74, 134, 1995/3).
Jak se množí DNA
Pro výrobu nových kopií DNA – replikaci – je klíčovou vlastností komplementarita (viz Molekuly na povel I. a II.). Tato vlastnost určuje, jak podle jednoho vlákna DNA vzniká vlákno nové, komplementární k původnímu: proti každému nukleotidu s adeninem je zařazen nukleotid s tyminem (a naopak) a proti každému nukleotidu s guaninem nukleotid s cytozinem (a taky naopak).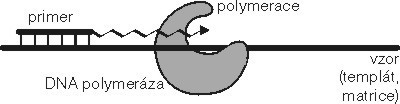 Proces tvorby nového vlákna se nazývá polymerace a provádí jej DNA polymeráza, což je většinou celý komplex enzymů a pomocných faktorů. Polymerace probíhá podle několika důležitých pravidel, z nichž jedno je do omrzení zmiňovaná komplementarita. Dalším pravidlem je, že výroba nového vlákna DNA podle vzoru nikdy nezačíná z ničeho, ale vždy je jenom prodlužováním již existující, třeba i velmi krátké molekuly. Tato krátká molekula se jmenuje primer a bez ní nemůže být syntéza nové DNA zahájena.
Proces tvorby nového vlákna se nazývá polymerace a provádí jej DNA polymeráza, což je většinou celý komplex enzymů a pomocných faktorů. Polymerace probíhá podle několika důležitých pravidel, z nichž jedno je do omrzení zmiňovaná komplementarita. Dalším pravidlem je, že výroba nového vlákna DNA podle vzoru nikdy nezačíná z ničeho, ale vždy je jenom prodlužováním již existující, třeba i velmi krátké molekuly. Tato krátká molekula se jmenuje primer a bez ní nemůže být syntéza nové DNA zahájena.
 Druhou zásadou polymerace je, že probíhá vždy jen v jednom směru. To zní trochu záhadně, ale vzpomeňte si na to, jak jsou ve vlákně DNA nukleotidy vzájemně navázané:
Druhou zásadou polymerace je, že probíhá vždy jen v jednom směru. To zní trochu záhadně, ale vzpomeňte si na to, jak jsou ve vlákně DNA nukleotidy vzájemně navázané:
 V nukleotidu má každý atom své číslo, což umožňuje těm, kteří systém číslování pochopili, orientaci v složité molekule (těm, kteří systém nepochopí, to naopak orientaci v podstatě znemožňuje). Uhlík v cukerné složce, na němž je navázán zbytek kyseliny fosforečné, má číslo 5´, uhlík, na němž je navázána skupina OH, má číslo 3´. Z toho vyplývá, že vlákno DNA je orientované, podle konvence má začátek 5´ a konec 3´ (chiliastičtí molekulární biologové ovšem říkají v obou případech konec: 5´ konec a 3´ konec). A teď pozor! Dvě vlákna ve dvoušroubovici DNA jsou nejenom komplementární, ale jsou také opačně orientovaná.
V nukleotidu má každý atom své číslo, což umožňuje těm, kteří systém číslování pochopili, orientaci v složité molekule (těm, kteří systém nepochopí, to naopak orientaci v podstatě znemožňuje). Uhlík v cukerné složce, na němž je navázán zbytek kyseliny fosforečné, má číslo 5´, uhlík, na němž je navázána skupina OH, má číslo 3´. Z toho vyplývá, že vlákno DNA je orientované, podle konvence má začátek 5´ a konec 3´ (chiliastičtí molekulární biologové ovšem říkají v obou případech konec: 5´ konec a 3´ konec). A teď pozor! Dvě vlákna ve dvoušroubovici DNA jsou nejenom komplementární, ale jsou také opačně orientovaná.
Pokud se vám to zdá komplikované, tak věřte tomu, že to nejhorší teprve přijde. DNA polymeráza, která podle vzoru prodlužuje na principu komplementarity nějaký primer, to dovede pouze jedním směrem, a to od 5´ ke 3´ konci. Všechny enzymy, které vytvářejí nukleové kyseliny, to dovedou pouze v tomto směru, to znamená, že na 3´OH skupinu předchozího nukleotidu napojí 5´ P skupinu dalšího nukleotidu. Jinak to nejde, jiné způsoby jsou zakázány, všechny polymerázy to dělají takhle. Přestože je to vlastně svým způsobem komplikace (například z toho vyplývají jisté topologické problémy při replikaci chromozomů, kterými vás ovšem nebudu zatěžovat), nedá se s tím nic dělat – vlákna v dvoušroubovici jsou opačně orientovaná a syntéza nového vlákna probíhá pouze prodlužováním primeru ve směru od 5´ ke 3´ konci.
Řetězová reakce
Všechny výše zmiňované detaily o syntéze nového vlákna jsou nezbytné k pochopení principu až podezřele jednoduché techniky polymerázové řetězové reakce. Představte si, že chcete namnožit do obrovského počtu kopií nějaký NOF, bez něhož nemůžete žít. Znáte pořadí nukleotidů koncových oblastí NOF nebo jeho okolí. Chemickou syntézou si necháte připravit dva primery, každý pro jedno vlákno DNA:
Smísíte DNA, která obsahuje NOF a bude sloužit jako templát (vzor) pro namnožení, s připravenými primery, a zahřejete na 96 °C. Pokud si dobře pamatujete, tak při této teplotě denaturuje, tedy rozpadá se na dvě samostatná vlákna, i ta nejpevnější dvoušroubovice. Potom celou směs ochladíte. Z vláken by se měla zase vytvořit dvouvlákna, ale dříve než se utvoří dvoušroubovice z dlouhých molekul DNA, nasednou na příslušná komplementární místa primery, které jste si přichystali, a zabrání opětnému vytvoření původních dvoušroubovic. Pokud k této směsi jednovláknových molekul DNA s nasednutými primery přidáte DNA polymerázu a prekurzory výroby DNA (nukleosidtrifosfáty), enzym zahájí prodlužování primerů podle templátu. Necháte nějakou dobu vyrábět nová vlákna, a pak směs znovu nemilosrdně zahřejete na 96 °C. Vzniklé dvouřetězce starých a nových vláken se rozpadnou, takže se ve směsi objeví zase spousta jednořetězců, v ideálním případě dvojnásobek výchozího počtu. Pak cyklus zopakujete. Ochladíte směs na teplotu optimální pro nasedání primerů, znovu spustíte polymeraci, znovu denaturujete, a tak pořád dokola. V každém cyklu vám vzniknou nová vlákna, která jsou na koncích „zaostřená“ primery. Jejich počet se v každém cyklu zdvojnásobuje (viz schéma). Cyklů můžete provést libovolně mnoho – třeba 30. Schválně si zkuste spočítat, kolik máte na konci nových molekul NOF!
Postup vypadá jednoduše, ale nelze při něm používat běžné DNA polymerázy např. z Escherichia coli, které mají optimální polymerační teplotu 37 °C a při teplotě nad 50 °C spolehlivě a nevratně stávkují. Pro polymerázovou řetězovou reakci (anglicky polymerase chain reaction, PCR, v české laboratorní hantýrce písíár) se používají polymerázy z termofilních mikroorganizmů, které tyto teploty bez problémů vydrží. Nejznámější a nejpoužívanější termostabilní polymeráza je Taq, která pochází z eubakterie Thermus aquaticus, méně známé, ale vcelku běžné jsou například polymerázy Tth (z eubakterie Thermus thermophillus) nebo Pwo (izolovaná z hypertermofilní archey Pyrococcus woesei). Nikdo nikdy nemohl odhadnout, k čemu se nám zkoumání takto bizarních organizmů může hodit – a vida, bez nich by PCR těžko dosáhla takového vzmachu (to je poznámka pro reptaly na základní výzkum). V současné době se také objevují rekombinantní – geneticky upravené – polymerázy, které mají (podle výrobců) optimalizované vlastnosti pro různé speciální postupy.
Všechno to (doufám) vypadá hrozně jednoduše, ale zas tak úplně snadné to není. Pro výsledek pokusu je totiž klíčové naplánovat optimální složení reakční směsi, v níž rozhoduje koncentrace templátové DNA, primerů i nezbytného pufru. A také sestavit nejvhodnější program, jako např. v automatické pračce, s tím rozdílem, že tam jsou programy praní připraveny od výrobců, zatímco pro PCR si termocykler (tak se ten přístroj jmenuje) programujete sami. Při tom je rozhodující zejména teplota nasedání primerů, která se odvozuje z jejich složení a velikosti. Pokud totiž určíte teplotu příliš nízkou, můžete získat nespecifické fragmenty (i z primerů, které nasedly na místa komplementární jen částečně). Pokud určíte teplotu naopak příliš vysokou, nemusíte dostat nic, protože primer nenasedne nikde.  A tak někdy přijdete o mnoho vlasů, přátel i iluzí, než se vám podaří vymyslit funkční program pro namnožení NOF polymerázovou řetězovou reakcí ( obr. 2 ).
A tak někdy přijdete o mnoho vlasů, přátel i iluzí, než se vám podaří vymyslit funkční program pro namnožení NOF polymerázovou řetězovou reakcí ( obr. 2 ).
A co s tím?
Nejdůležitější na polymerázové řetězové reakci je, že nám umožňuje získat velké množství DNA i z několika málo buněk. Zanechá-li pachatel na místě činu jediný vlas vytržený i s kořínkem, je to jako by tam nechal svůj kartotéční lístek s otisky prstů (jiná věc je pak zjistit, u koho ty příslušné otisky hledat). Z ubohých zbytků rozlámané a poškozené DNA mumií egyptských faraonů byly namnoženy úseky genů, které porovnáním s týmiž úseky našich současníků vydávají neuvěřitelná svědectví o lidských rasách, jejich historii a evoluci. Důležitou vlastností PCR ovšem není jenom možnost získat úseky DNA z několika málo buněk. Polymerázová řetězová reakce je také velmi specifická – namnoží se pouze úsek DNA sevřený mezi primery, které jsme připravili. Díky tomu můžeme rozeznat i jedinou určitou buňku (přesněji její DNA) mezi stovkami jiných – a například velmi brzy rozhodnout, zda se transplantace kostní dřeně zdařila nebo ne.K polymerázové řetězové reakci lze jako templát použít také RNA. Tato takzvaná reverzní PCR (RT-PCR) využívá reverzní transkriptázu, která přepisuje RNA na DNA. Díky RT-PCR jsme získali citlivou metodu k určování aktivních genů v buňce – tedy těch genů, z nichž se vytváří mRNA, a to i ve velmi malém množství.
 Důležité je využití technik odvozených od PCR k identifikaci jedinců i druhů. Nejběžnější je technika RAPD (random amplified polymorphic DNA), při které se pro PCR používá pouze jeden velmi krátký primer. Předpokládáme, že tento krátký primer najde v testovaném genomu náhodně několik komplementárních míst, k nimž nasedne, a úseky těmito primery sevřené namnožíme. Je jasné, že v každém organizmu budou komplementární místa různá, a tudíž i vzniklé fragmenty budou různě dlouhé a bude jich různý počet. Fragmenty získané ze stejného primeru za stejných podmínek u různých organizmů pak můžeme porovnat (obr. 3) elektroforetickou analýzou.
Důležité je využití technik odvozených od PCR k identifikaci jedinců i druhů. Nejběžnější je technika RAPD (random amplified polymorphic DNA), při které se pro PCR používá pouze jeden velmi krátký primer. Předpokládáme, že tento krátký primer najde v testovaném genomu náhodně několik komplementárních míst, k nimž nasedne, a úseky těmito primery sevřené namnožíme. Je jasné, že v každém organizmu budou komplementární místa různá, a tudíž i vzniklé fragmenty budou různě dlouhé a bude jich různý počet. Fragmenty získané ze stejného primeru za stejných podmínek u různých organizmů pak můžeme porovnat (obr. 3) elektroforetickou analýzou.
To je ovšem jen zlomek možností, které nám PCR poskytuje. V současné době je to jedna z nejúčinnějších a nejvíce se rozvíjejících metod molekulární biologie, a tak není divu, že její objevitelé byli r. 1993 oceněni Nobelovou cenou za chemii (viz Vesmír 73, 8, 1994/1).
Několik čísel o DNA
Je DNA v buňce hodně, nebo málo? Kolik jí molekulární biolog vlastně potřebuje k práci?
Molekulární hmotnost DNA se odhaduje na 333násobek počtu nukleotidů (při průměrném zastoupení jednotlivých bází). 1 g fragmentu dvouřetězcové DNA o velikosti 1000 bp (párů bází) činí 1,52 pmol, 1 pmol stejně velkého fragmentu váží 666 ng. Genom bakterie Escherichia coli má více než 4 × 106 bp, haploidní genom Saccharomyces cerevisiae odpovídá zhruba 1,5 × 107 bp. V jedné buňce pánů tvorstva se nachází 4 × 109 bp. Na elektroforéze pohodlně vidíme nanogramy (10–6 g) DNA. V jedné kvasinkové buňce je asi 2,5 × 10–14 g DNA, pro jednu elektroforetickou analýzu kvasinek tedy musíme izolovat DNA z 107 až 108 buněk. Průměrný savčí gen má délku 3 kbp, což činí asi 0,000075 % celého genomu. Hybridizací jsme schopni detegovat (máme-li kvalitní sondu) zhruba desetiny (až setiny) pikogramu. Potřebujeme tedy DNA nejméně z 105 až 106 buněk (jedna buňka obsahuje 6,7 × 10–3 pg DNA), abychom se mohli spolehnout na to, že hledaný úsek DNA nalezneme.
Nedostatky PCR
Každý, kdo se s polymerázovou řetězovou reakcí setkal, má tendenci vynášet ji do nebe, a budiž řečeno, že právem. Ale samozřejmě i tato technika má své problémy.
- Jednou komplikací je právě její účinnost a specifita. Stačí i třeba minimální kontaminace nějakou cizí DNA, například zanesení DNA do roztoku používaných primerů, a výsledky reakce ztrácejí jakoukoliv výpovědní hodnotu. V laboratořích specializovaných na diagnostiku s využitím PCR nebo v laboratořích kriminalistů znají řadu postupů jak riziko kontaminace snížit. V mnoha případech jde totiž do slova a do písmene o život.
- Druhou vadou PCR je nepřesnost používaných enzymů. Termostabilní polymerázy pracují v extrémních podmínkách a poměrně často (podstatně častěji než ostatní DNA polymerázy) zařazují do řetězce chybné nukleotidy. Pro přesné syntézy konkrétních úseků genů je proto třeba věnovat dostatečnou pozornost kontrole výsledku.
Rozhodně však metoda polymerázové řetězové reakce podnikla triumfální tažení laboratořemi a nezdá se, že by někdy v budoucnu musela získané pozice vyklízet.
Ke stažení
 článek ve formátu PDF [368,16 kB]
článek ve formátu PDF [368,16 kB]
O autorovi
Zuzana Storchová
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
