













Molekuly na povel I.
V novinách i nejrůznějších časopisech se často dočítáme o úžasných úspěších molekulární biologie, genetiky, biotechnologií a dalších podobných věd. S překvapením zjišťujeme, že byly objeveny geny paměti, alkoholizmu, schizofrenie, schopnosti učení, a je nám jasné, že za chvíli bude objeven i gen optimizmu, charizmatu či sklonů k prostopášnosti. Vědci zřejmě zcela nekontrolovaně v laboratořích klonují každé zvíře, které jim přijde pod ruku, vyrábějí chimérická zvířátka s ušima na břiše a oživují dávno vymřelé organizmy.
Skutečnost je daleko prozaičtější a zároveň mnohem zajímavější. Molekulární biolog je většinou úplně obyčejný člověk, který ovšem zkoumá něco neviditelného, něco, o jehož existenci a podobě se dovídá pouze prostřednictvím řady detekčních technik. Jeho práce je závislá na používaných metodách, které jsou většinou nesmírně složité a technicky, časově i finančně náročné. Za celý svůj život i velmi dobrý molekulární biolog dokonale zvládne jen některé z existujících technik, jejichž spektrum se navíc neustále rozšiřuje.
 Ačkoliv prováděcí předpisy metod molekulární biologie jsou většinou k uzoufání komplikované, princip používaných metod je průzračně jednoduchý. Zkusme se tedy podívat, co vlastně ti molekulárníci v laboratořích provádějí. Není třeba se bát, že to obyčejný smrtelník nedokáže pochopit. Už to, že jste dočetli text až sem, svědčí o vaší dostatečné kvalifikaci – molekulární biolog totiž ke své práci potřebuje znát pouze 4 písmena: A, G, C a T.
Ačkoliv prováděcí předpisy metod molekulární biologie jsou většinou k uzoufání komplikované, princip používaných metod je průzračně jednoduchý. Zkusme se tedy podívat, co vlastně ti molekulárníci v laboratořích provádějí. Není třeba se bát, že to obyčejný smrtelník nedokáže pochopit. Už to, že jste dočetli text až sem, svědčí o vaší dostatečné kvalifikaci – molekulární biolog totiž ke své práci potřebuje znát pouze 4 písmena: A, G, C a T.
Několik užitečných vlastností DNA
Objektem zájmu molekulárních biologů jsou bílkoviny a nukleové kyseliny (nukleové znamená jaderné a ony se skutečně vyskytují v jádře, i když nejenom tam). Nechme bílkoviny prozatím stranou a soustřeďme svůj zájem na nukleové kyseliny, které se v organizmech vyskytují ve dvou podobách. Obě mají podobnou strukturu, složení i vlastnosti a liší se jen v drobných, ale podstatných detailech. Kostra ribonukleové kyseliny (RNA) je tvořena cukrem ribózou, zatímco u deoxyribonukleové kyseliny najdeme kupodivu deoxyribózu. Drobný chemický rozdíl mezi těmito dvěma molekulami způsbuje, že RNA je mnohem reaktivnější a méně stabilní než DNA. Proto je jako trvalý dědičný materiál v buňce používána DNA, zatímco RNA slouží jako poslíček na přenos a zprostředkování údajů z DNA. Jednotlivé molekuly cukru jsou u nukleových kyselin spojeny pomocí fosfátového zbytku a na každý cukr je navázána jedna molekula dusíkaté sloučeniny – báze. Základní jsou čtyři báze, označované jako A – adenin, G – guanin, C – cytozin a T – tymin, které najdeme u DNA. Nicméně znalost těchto čtyř písmen molekulárnímu biologovi přece jenom nestačí. V RNA je totiž místo tyminu báze uracil – U, která se ovšem chová jako úplně prachobyčejný tymin. Cukr s navázanou bází a fosfátem se nazývá nukleotid, a každá molekula nukleových kyselin je tedy vlastně polynukleotid – prostě moc nukleotidů za sebou. Pořadí nukleotidů v molekule je onou závažnou genetickou informací, která určuje vaši plešatost, barvu očí, počet rukou a kdovíco ještě.Jednotlivé nukleotidy se mohou spojovat nejenom do řad za sebou pomocí fosfátů, ale také se mohou chytit napřaženými ručičkami bází. Tato vazba je slabá a funguje dobře pouze tehdy, když je těch ručiček dostatek. Jak známe i z vlastního života, nemůže se ovšem za ručičku držet každý s každým. Pro nukleové kyseliny platí pravidlo, že se drží jen adenin s tyminem (nebo uracilem) a druhou dvojici tvoří jen cytozin s guaninem. Dodržování tohoto pravidla je pojištěno docela jednoduše – adenin, tymin a uracil mají dvě ručičky a pasují tedy pouze k sobě, zatímco trojručičkový guanin má na vybranou jenom cytozin, taktéž s třemi ručkami. A protože tři ruce jsou více než dvě, je vazba mezi guaninem a cytozinem o něco silnější.
 Spojení molekul nukleových kyselin do dvouřetězce je možné pouze tam, kde jsou proti sobě ty báze, které se umějí držet za ručičky. A protože je vazba mezi bázemi poměrně slabá, nestačí k udržení molekul jenom jedna nebo dvě dvojice.
Spojení molekul nukleových kyselin do dvouřetězce je možné pouze tam, kde jsou proti sobě ty báze, které se umějí držet za ručičky. A protože je vazba mezi bázemi poměrně slabá, nestačí k udržení molekul jenom jedna nebo dvě dvojice.
Této vlastnosti řetězců se říká komplementarita (doplňkovost) a je jednou z nejzajímavějších vlastností nukleových kyselin – umožňuje totiž jejich rozmnožování.
Jen díky tomu, že před každým rozdělením buňky na dvě nové dojde ke zdvojení genetického materiálu, může se dědičná informace dědit.
Stříhání a štípání
Opravdový rozmach molekulární biologie, tedy přesněji kejklů s nukleovými kyselinami, byl umožněn objevem restrikčních endonukleáz. Tito vnitromolekuloví střihači nukleových kyselin (téměř přesný překlad jejich názvu) pocházejí z bakterií, kde mají zcela speciální a mimochodem velmi zajímavou úlohu – rozstříhají každou cizí nukleovou kyselinu, která náhodou vleze do buňky. Během 70. let bylo objeveno obrovské množství restriktáz z nejrůznějších bakterií. Nejzajímavější jsou ovšem restriktázy II. typu, které dovedou stříhat DNA v úplně přesném místě. Vždycky v molekule najdou stejné místo, kde přeruší vazbu mezi cukrem a fosfátem a zanechají po sobě pouze dva volné konce. Rozeznávané místo ovšem nemůže být jen tak ledajaké. Vždy má jednu zajímavou vlastnost – čteme-li ho protisměrně na obou řetězcích, zjistíme, že má stejné pořadí bází. Takové oblasti dvoušroubovice DNA říkáme palindrom. Jen takové místo dovedou restriktázy II. typu najít. Takto například vypadá místo a zásah restriktázy EcoRI, která byla izolována z bakterie Escherichia coli, kmene R.
molekulární biologie, tedy přesněji kejklů s nukleovými kyselinami, byl umožněn objevem restrikčních endonukleáz. Tito vnitromolekuloví střihači nukleových kyselin (téměř přesný překlad jejich názvu) pocházejí z bakterií, kde mají zcela speciální a mimochodem velmi zajímavou úlohu – rozstříhají každou cizí nukleovou kyselinu, která náhodou vleze do buňky. Během 70. let bylo objeveno obrovské množství restriktáz z nejrůznějších bakterií. Nejzajímavější jsou ovšem restriktázy II. typu, které dovedou stříhat DNA v úplně přesném místě. Vždycky v molekule najdou stejné místo, kde přeruší vazbu mezi cukrem a fosfátem a zanechají po sobě pouze dva volné konce. Rozeznávané místo ovšem nemůže být jen tak ledajaké. Vždy má jednu zajímavou vlastnost – čteme-li ho protisměrně na obou řetězcích, zjistíme, že má stejné pořadí bází. Takové oblasti dvoušroubovice DNA říkáme palindrom. Jen takové místo dovedou restriktázy II. typu najít. Takto například vypadá místo a zásah restriktázy EcoRI, která byla izolována z bakterie Escherichia coli, kmene R.
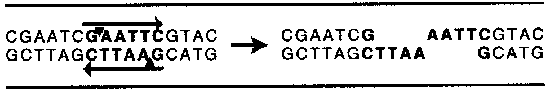
To, co po ní zůstalo, se jmenuje lepivé konce, protože se to může přilepit k jakékoliv jednořetězcové molekule se stejným pořadím nukleotidů.
Jiná restriktáza BstUI z mikroorganizmu Bacillus stearothermophillus, kmene U, po sobě zanechá jiné stopy. Vzniklý konec se jmenuje tupý a myslím, že je zřejmé proč.

Dnes známe už řadu restriktáz, které štěpí na nejrůznějších místech. Díky jejich činnosti můžeme rozsekat dlouhé molekuly DNA na různé kousky, které mají konce o známém pořadí nukleotidů. K čemu nám ale takové kousíčky molekul vlastně jsou?
Spojování a lepení
Restriktázy by samozřejmě nikdy nedosáhly takové slávy, kdyby neexistovaly jejich protějšky – ligázy, které umějí kousky molekul zase spojit. Restriktáza rozbije vazbu mezi cukrem a fosfátem, ligáza ji zase spojí. Musíme jí k tomu samozřejmě vytvořit vhodné podmínky a dodat energii. Hlavně ale musíme spojovat molekuly, jejichž konce k sobě patří. Lze tedy spojit pouze kousky DNA vzniklé štěpením restriktázou EcoRI, a nikoliv vzniklé po štěpení EcoRI a BstUI. Jinak k sobě naštěpené kousíčky prostě nepasují.
To ovšem neplatí jenom pro spojování lepivých a tupých konců, ale i pro spojování dvou lepivých konců s různým pořadím nukleotidů. Jakmile přečnívající ocásky nejsou komplementární, tedy pořadí bází sobě neodpovídá, nemohou se báze lepivých konců chytit za ručičky a ligáza nemá co spojovat.

Stejně jako výše zmiňované restriktázy, i ligázy jsou zcela běžnými enzymy buněk, tentokrát dokonce nejenom bakteriálních. Bez nich by vůbec celá ta mašinérie DNA v buňce nefungovala. My jsme je pouze poznali, porozuměli jejich činnosti a naučili se je využívat.
K čemu to celé vlastně je?
Díky restriktázám můžeme jakoukoliv DNA naštěpit na menší molekuly s definovanými konci. Díky ligázám spolu dovedeme kousky se shodnými konci spojit. Trik spočívá v tom, že můžeme spojit jakoukoliv DNA z jakéhokoliv organizmu s molekulou pocházející z jakéhokoliv organizmu jiného. Stačí, když mají spojované kousky stejné konce. A tak můžeme kousek molekuly DNA, který v člověčích buňkách vydává instrukce k výrobě inzulinu, spojit s kouskem DNA, který umožní jeho udržení v bakteriích. Když pak takovouto pospojovanou, chimérickou, rekombinantní DNA, vzniklou z kousků molekul dvou různých organizmů, přeneseme do bakterií (a ještě s tím provedeme nějaké další složité kejkle), můžeme docílit toho, že nebohé bakterie začnou ve velkém vyrábět inzulin. No, a hádejte, jak se dneska (nejenom) inzulin vyrábí.Štěpení a spojování molekul DNA je základní metodou molekulární biologie a představuje vstupní bránu k dalším technikám. Projděte si ještě jednou celý článek – není to jednoduché? 1)
Poznámky
Ke stažení
 Článek ve formátu PDF [107,54 kB]
Článek ve formátu PDF [107,54 kB]
O autorovi
Zuzana Storchová
Doporučujeme

Když bahno teče jako ledovec

Příliš otevřené dveře

