









Metabolismus a velikost těla v novém tisíciletí
Jednou z největších událostí v biologii konce minulého tisíciletí byl vznik metabolické teorie biologie (viz Vesmír 83, 508, 2004/9). Díky spolupráci biologů a fyziků v Santa Fe Institute se podařilo navrhnout vysvětlení sedmdesát let staré záhady, proč intenzita metabolismu u živočichů roste s hmotností těla podle mocninné zákonitosti s exponentem ¾ (rámeček). Jim Brown, Brian Enquist a Geoffrey West tvrdili, že za vše jsou odpovědné větvící se rozvodné systémy (cévní soustava, plíce, vylučovací soustava), jejichž rozsah nutně roste s velikostí těla tak, že dvakrát větší zvíře nemůže mít dvakrát intenzivnější metabolismus (čemuž by odpovídalo škálování s exponentem 1), ale o něco menší. Při optimálním uspořádání nelze dosáhnout vyššího exponentu, než jsou právě tři čtvrtiny. Takový exponent se pak nacházel ve škálování skoro všech biologických procesů a mělo se za to, že je univerzální a platí pro všechny organismy, včetně jednobuněčných. U těch ovšem neexistují žádné větvící se rozvodné systémy.
Během prvního desetiletí nového tisíciletí byla metabolická teorie aplikována na nejrůznější problémy a systémy, včetně lidských společností. Zároveň ale jiní autoři její teoretická východiska i empirické závěry zpochybňovali. Někteří tvrdili, že onen tříčtvrtinový exponent není zdaleka univerzální, a navrhovali alternativní vysvětlení pozorovaného škálování (založená třeba na předpokladu, že škálovací koeficient může libovolně kolísat mezi jedničkou, která odpovídá lineárnímu vztahu, a dvěma třetinami, které by souvisely s nárůstem povrchu s rostoucím objemem). Dostupná data ale většinou nedovolovala rozhodnout, kdo má pravdu, poněvadž chyba měření byla relativně velká a různé statistické metody docházely na základě různých předpokladů k různým závěrům.
Modifikace metabolické teorie
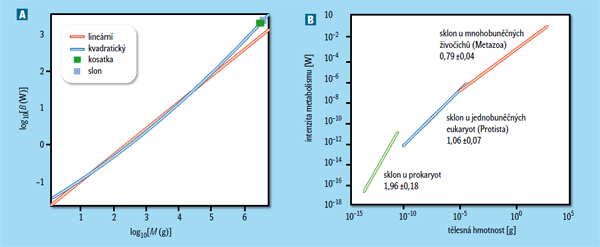 Letos se objevilo několik přelomových studií, které celou problematiku posunuly na novou úroveň. Ukázalo se například, že srovnání různě velkých jedinců v rámci jednoho druhu vede k jinému škálování metabolismu, než když mezi sebou srovnáváme různé druhy. Vnitrodruhové škálování můžeme dobře studovat u ryb, kde i v rámci druhu existují obrovské rozdíly ve velikosti. I zde nalezneme tříčtvrtinový koeficient, jenže jen u těch druhů, které mají obecně intenzivní metabolismus spojený s aktivním způsobem života. U „líných“ ryb s pomalým metabolismem, například ryb žijících u dna, roste jeho intenzita s velikostí téměř lineárně.1) Rozpor s metabolickou teorií však není tak příkrý, jak by se zdálo; je možné, že u ryb s aktivním metabolismem jsou skutečně limitující ony větvené rozvodné systémy, zatímco u pomalých ryb stačí rozvodné systémy pohodlně dodávat buňkám všechny živiny (a odebírat odpadní produkty), a tak je jejich metabolismus úměrný pouze celkovému počtu buněk, a tedy roste (skoro) lineárně s velikostí těla.
Letos se objevilo několik přelomových studií, které celou problematiku posunuly na novou úroveň. Ukázalo se například, že srovnání různě velkých jedinců v rámci jednoho druhu vede k jinému škálování metabolismu, než když mezi sebou srovnáváme různé druhy. Vnitrodruhové škálování můžeme dobře studovat u ryb, kde i v rámci druhu existují obrovské rozdíly ve velikosti. I zde nalezneme tříčtvrtinový koeficient, jenže jen u těch druhů, které mají obecně intenzivní metabolismus spojený s aktivním způsobem života. U „líných“ ryb s pomalým metabolismem, například ryb žijících u dna, roste jeho intenzita s velikostí téměř lineárně.1) Rozpor s metabolickou teorií však není tak příkrý, jak by se zdálo; je možné, že u ryb s aktivním metabolismem jsou skutečně limitující ony větvené rozvodné systémy, zatímco u pomalých ryb stačí rozvodné systémy pohodlně dodávat buňkám všechny živiny (a odebírat odpadní produkty), a tak je jejich metabolismus úměrný pouze celkovému počtu buněk, a tedy roste (skoro) lineárně s velikostí těla.
Jenže složitější je to i u savců, což je skupina, u níž byl tříčtvrtinový koeficient poprvé přesvědčivě doložen. Vyneseme-li proti sobě hmotnost a intenzitu metabolismu v logaritmických osách, detailní analýza ukáže, že vztah není přesně lineární, jak by předpokládala ona mocninná závislost (rámeček), ale mírně se zakřivuje směrem nahoru2) (obr. 1A). Jako by větší savci měli vyšší škálovací koeficient než menší savci. Autoři ukazují, že pozorovanou odchylku lze vysvětlit poměrně drobnou modifikací původního modelu. Předpokládáme-li, že postupně se větvící rozvodné systémy mají jen několik úrovní větvení, poněvadž každé zvíře má jen omezenou velikost, a že na určité úrovni větvení se mění povaha toku látek od pulzujícího k rovnoměrnému, skutečně získáme mírně zakřivený vztah.
Tím ale příběh nekončí. Nick Isaac a Chris Carbone detailně analyzovali vztah hmotnosti těla a metabolismu u obrovského množství živočišných druhů a došli k závěru, že škálovací koeficient je u různých taxonů různý, i když v průměru vycházejí ony tři čtvrtiny.3) Ty tedy představují spíše jakousi centrální tendenci s více či méně výraznými odchylkami. Z toho zároveň plyne, že nemá smysl analyzovat vztah mezi hmotností a metabolismem pro příliš široce vymezené skupiny, protože ty mohou zahrnovat několik podskupin s odlišným škálováním. Tím lze možná vysvětlit i ono zakřivení pozorované u savců; autoři totiž ukázali, že nejtěsnější vztahy lze pozorovat v rámci jednotlivých řádů (jakkoli je jasné, že tato hierarchická úroveň je do značné míry arbitrární), zatímco vztah pro větší taxonomické skupiny, jako jsou třídy, je dán kombinací vztahů pro jednotlivé řády s mírně odlišnými koeficienty. Variabilitu koeficientů mezi různými taxony ovšem zatím nelze vysvětlit na základě žádné dosud navržené teorie.
Škálování a evoluce komplexity
U jednobuněčných je ale všechno jinak. Nejnovější analýzy ukázaly, že metabolismus prokaryotních organismů (bakterie a archea) roste dokonce rychleji než jejich velikost těla, škálovací koeficient je blízký dvěma (což by představovalo kvadratickou závislost), zatímco protisti (jednobuněčné eukaryotní organismy) vykazují lineární závislost4) (obr. 1B). Autoři spekulují, že změny metabolického škálování souvisí s dvěma základními evolučními přechody: vznikem eukaryotní buňky a vznikem mnohobuněčnosti. U prokaryot je podle nich metabolismus omezen především množstvím genů produkujících proteiny, které se účastní metabolických reakcí. S velikostí buňky roste totiž velikost genomu, a ta u prokaryot poměrně dobře odpovídá počtu genů kódujících proteiny, protože (na rozdíl od eukaryot) mají málo nekódujících sekvencí. Když jsou ale prokaryotní buňky příliš velké, stane se relativně malý povrch buňky, na němž metabolické reakce probíhají, omezujícím faktorem. V té chvíli začnou být konkurenčně zvýhodněny eukaryotní buňky, u nichž metabolické pochody probíhají na membránách mitochondrií, organel vzniklých endosymbiotickou adopcí prokaryotních buněk. Metabolismus zde patrně roste lineárně s počtem mitochondrií, a tedy s velikostí celé buňky. Jenže když jsou buňky příliš velké, nastává zase problém s účinnou dopravou látek do mitochondrií uvnitř buňky. V této velikostní kategorii je výhodnější být mnohobuněčný s menšími buňkami – jenže k těm buňkám je třeba příslušné látky nějak dostat, a čím jich je více, tím rozsáhlejší jsou příslušné rozvodné systémy, což vede k našemu známému tříčtvrtinovému škálování.
Je to pěkná pohádka. Metabolismus je klíčovým biologickým procesem, a tak je možné, že jsou základní evoluční přechody spojeny s překonáním limitů omezujících jeho intenzitu a zároveň s nastolením nových omezení. Něco podobného by mohlo vysvětlit i výše zmiňovanou různorodost škálovacího koeficientu mezi různými taxony, poněvadž každý taxon čelí trochu jiným omezením. Jaká ta omezení jsou a co přesně způsobuje variabilitu v naměřených koeficientech, pořád ještě nevíme. Každopádně se zdá, že u mnohobuněčných organismů nebyl tříčtvrtinový koeficient naměřen jen náhodou, ale jde o kanonickou hodnotu vyjadřující základní dimenzionální omezení jejich světa. Odchylky pak představují biologicky zajímavou informaci, specifickou pro jednotlivé taxonomické skupiny.
Poznámky
1) Killien S. S., Atkinson D., Glazier D. S.: The intraspecific scaling of metabolic rate with body mass in fishes depends on lifestyle and temperature, Ecology Letters 13, 184–193, 2010.
2) Kolokotrones T., Savage V., Deeds E. J., Fontana W.: Curvature in metabolic scaling, Nature 464, 753–756, 2010.
3) Isaac N. J. B., Carbone C.: Why are metabolic scaling exponents so controversial? Quantifying variance and testing hypotheses, Ecology Letters 13, 728–735, 2010.
4) DeLong J. P., Okie J. G., Moses M. E., Sibly R. M., Brown J. H.: Shifts in metabolic scaling, production, and efficiency across major evolutionary transitions of life. PNAS, in review.
5) Starostová Z., Kratochvíl L., Frynta D.: Dwarf and giant geckos from the cellular perspective: the bigger the animal, the bigger its erythrocytes? Functional Ecology 19, 744–749, 2005.
6) Starostová Z., Kubička L., Konarzewski M., Kozłowski J., Kratochvíl L.: Cell size but not genome size affects scaling of metabolic rate in eyelid geckos. American Naturalist 174, E100–E105, 2009.
7) Kozłowski J., Czarnołeski M., François-Krassowska A., Maciak S., Pis T.: Cell size is positively correlated between different tissues in passerine birds and amphibians, but not necessarily in mammals. Biol. Lett., in press, 2010.
 Vyneseme-li proti sobě hmotnost těla a intenzitu metabolismu (ta se měří třeba spotřebou kyslíku za jednotku času, ale lze ji udávat ve wattech, poněvadž jde vlastně o příkon), dostaneme křivku, jejíž růst se postupně zpomaluje (a). Učiníme-li to tak, že jsou obě osy logaritmické (b), získáme závislost, kterou lze velmi dobře proložit přímkou s určitým sklonem – jedna veličina škáluje s druhou. Vzhledem k tomu, že rovnice přímky je y = ax + b, kde a je onen sklon, můžeme rovnici vztahu intenzity metabolismu a hmotnosti vyjádřit jako logB = a.logM + logB0, kde B je intenzita metabolismu, M je hmotnost a logB0 je průsečík dané přímky se svislou osou. Po odlogaritmování dostaneme závislost B = B0.Ma, jde tedy o mocninnou funkci, kde sklon přímky v logaritmickém měřítku a se stává exponentem; hovoříme o škálovacím koeficientu. Ten bývá typicky nižší než 1 (a velmi často blízký třem čtvrtinám) a v takovém případě roste v nelogaritmickém měřítku metabolismus s hmotností čím dál pomaleji, jak je vidět z obrázku A.
Vyneseme-li proti sobě hmotnost těla a intenzitu metabolismu (ta se měří třeba spotřebou kyslíku za jednotku času, ale lze ji udávat ve wattech, poněvadž jde vlastně o příkon), dostaneme křivku, jejíž růst se postupně zpomaluje (a). Učiníme-li to tak, že jsou obě osy logaritmické (b), získáme závislost, kterou lze velmi dobře proložit přímkou s určitým sklonem – jedna veličina škáluje s druhou. Vzhledem k tomu, že rovnice přímky je y = ax + b, kde a je onen sklon, můžeme rovnici vztahu intenzity metabolismu a hmotnosti vyjádřit jako logB = a.logM + logB0, kde B je intenzita metabolismu, M je hmotnost a logB0 je průsečík dané přímky se svislou osou. Po odlogaritmování dostaneme závislost B = B0.Ma, jde tedy o mocninnou funkci, kde sklon přímky v logaritmickém měřítku a se stává exponentem; hovoříme o škálovacím koeficientu. Ten bývá typicky nižší než 1 (a velmi často blízký třem čtvrtinám) a v takovém případě roste v nelogaritmickém měřítku metabolismus s hmotností čím dál pomaleji, jak je vidět z obrázku A.
Je škálování metabolismu ovlivněno velikostí buněk?
 Velikost těla se může v podstatě měnit třemi způsoby a jejich kombinacemi: změnou v množství mezibuněčné hmoty, změnou v počtu buněk a změnou velikosti buněk. Každý ze způsobů přináší jiné morfologické a fyziologické změny. Z malého počtu velkých buněk se například nedá postavit komplexní orgán (zkuste si to s kostičkami Lega!) a malí živočichové s velkými buňkami – třeba mločíci – tak musí mít nutně zjednodušenou tělní stavbu, například málo strukturovaný mozek. Mechanismus změny ve velikosti těla by ale měl ovlivňovat i škálování celkového metabolismu. Změna velikosti nárůstem metabolicky spíše inertní tkáně (např. kostí) se jistě promítne do škálování intenzity metabolismu. Podobně může působit i změna ve velikosti nebo v počtu buněk. Předpokládá se, že větší buňky mají relativně menší povrch vzhledem k objemu, a tedy i relativně menší plochu buněčných membrán. A právě udržení integrity buněčných membrán je energeticky nesmírně náročné. Velké buňky by tedy neměly umožnit velký výkon, měly by ale být energeticky šetrnější. Mění se však velikost buněk systematicky s velikostí těla? To by mohlo vysvětlit část variability v rozdílech v relativní intenzitě metabolismu mezi malými a velkými organismy. Pro zodpovězení této otázky jsme si vybrali skupinu gekončíků. Zástupci této čeledi jsou si morfologicky a ekologicky podobní, přitom se však podstatně liší ve velikosti těla. Ukázali jsme, že mezi druhy gekončíků velikost červených krvinek roste s průměrnou velikostí těla5) a negativně koreluje s intenzitou metabolismu vztaženou na jednotku hmotnosti.6) Červené krvinky slouží k rozvodu kyslíku po těle, měly by tedy být zodpovědné aspoň za část variability v intenzitě metabolismu. Jistě lze namítnout, že červené krvinky představují jen jeden, a to vysoce specializovaný typ buněk. Nedávná práce však ukázala, že aspoň mezi druhy ptáků a obojživelníků spolu korelují i velikosti buněk různých typů z nejrůznějších tkání.7) Změny ve velikosti buněk tak mohou představovat důležitý faktor ovlivňující nejrůznější ekofyziologické vlastnosti a měly by být brány v úvahu i při vysvětlování variability škálovacích koeficientů mezi taxony.
Velikost těla se může v podstatě měnit třemi způsoby a jejich kombinacemi: změnou v množství mezibuněčné hmoty, změnou v počtu buněk a změnou velikosti buněk. Každý ze způsobů přináší jiné morfologické a fyziologické změny. Z malého počtu velkých buněk se například nedá postavit komplexní orgán (zkuste si to s kostičkami Lega!) a malí živočichové s velkými buňkami – třeba mločíci – tak musí mít nutně zjednodušenou tělní stavbu, například málo strukturovaný mozek. Mechanismus změny ve velikosti těla by ale měl ovlivňovat i škálování celkového metabolismu. Změna velikosti nárůstem metabolicky spíše inertní tkáně (např. kostí) se jistě promítne do škálování intenzity metabolismu. Podobně může působit i změna ve velikosti nebo v počtu buněk. Předpokládá se, že větší buňky mají relativně menší povrch vzhledem k objemu, a tedy i relativně menší plochu buněčných membrán. A právě udržení integrity buněčných membrán je energeticky nesmírně náročné. Velké buňky by tedy neměly umožnit velký výkon, měly by ale být energeticky šetrnější. Mění se však velikost buněk systematicky s velikostí těla? To by mohlo vysvětlit část variability v rozdílech v relativní intenzitě metabolismu mezi malými a velkými organismy. Pro zodpovězení této otázky jsme si vybrali skupinu gekončíků. Zástupci této čeledi jsou si morfologicky a ekologicky podobní, přitom se však podstatně liší ve velikosti těla. Ukázali jsme, že mezi druhy gekončíků velikost červených krvinek roste s průměrnou velikostí těla5) a negativně koreluje s intenzitou metabolismu vztaženou na jednotku hmotnosti.6) Červené krvinky slouží k rozvodu kyslíku po těle, měly by tedy být zodpovědné aspoň za část variability v intenzitě metabolismu. Jistě lze namítnout, že červené krvinky představují jen jeden, a to vysoce specializovaný typ buněk. Nedávná práce však ukázala, že aspoň mezi druhy ptáků a obojživelníků spolu korelují i velikosti buněk různých typů z nejrůznějších tkání.7) Změny ve velikosti buněk tak mohou představovat důležitý faktor ovlivňující nejrůznější ekofyziologické vlastnosti a měly by být brány v úvahu i při vysvětlování variability škálovacích koeficientů mezi taxony.
Lukáš Kratochvíl
Zuzana Starostová
Ke stažení
 článek ve formátu pdf [1,01 MB]
článek ve formátu pdf [1,01 MB]
O autorovi
David Storch

Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
