









Metabolická teorie biologie aneb Nová teorie všeho (živého)?
| 16. 9. 2004Biologové a fyzikové z Nového Mexika aspirují na vytvoření nové teorie, která vychází ze základních principů fyziky, chemie a biologie, a měla by postihnout většinu rozmanitosti a fungování organizmů.
 Nejnápadnější vlastností života je jeho rozmanitost. Množství způsobů, jimiž organizmy čelí nástrahám prostředí, je tak ohromné, až se zdá, že pokusy nějak se v tom vyznat musí nutně selhat. Každá skupina rostlin nebo živočichů totiž dostala do vínku jiná omezení, která určují, jak se budou jejich životní strategie vyvíjet. A tak dosud jediná úspěšná obecná biologie je ta, která se zabývá molekulami a jejich fungováním v buňce, poněvadž to je to jediné, co mají všechny organizmy více či méně společné. Jistěže v biologii existují i jiné obecné zákonitosti – třeba princip přirozeného výběru nebo Mendelovy zákony dědičnosti – jenže ty jsou zase až příliš obecné na to, aby nám mohly poskytnout nějaké předpovědi týkající se rozdílů a shod ve způsobu života různých druhů organizmů.
Nejnápadnější vlastností života je jeho rozmanitost. Množství způsobů, jimiž organizmy čelí nástrahám prostředí, je tak ohromné, až se zdá, že pokusy nějak se v tom vyznat musí nutně selhat. Každá skupina rostlin nebo živočichů totiž dostala do vínku jiná omezení, která určují, jak se budou jejich životní strategie vyvíjet. A tak dosud jediná úspěšná obecná biologie je ta, která se zabývá molekulami a jejich fungováním v buňce, poněvadž to je to jediné, co mají všechny organizmy více či méně společné. Jistěže v biologii existují i jiné obecné zákonitosti – třeba princip přirozeného výběru nebo Mendelovy zákony dědičnosti – jenže ty jsou zase až příliš obecné na to, aby nám mohly poskytnout nějaké předpovědi týkající se rozdílů a shod ve způsobu života různých druhů organizmů.
 Na druhou stranu víme, že různé vlastnosti organizmů spolu nějak souvisejí. Například větší organizmy se často dožívají vyššího věku, ale mívají méně potomků než ty menší. Takových vzájemných souvislostí známe spoustu, donedávna to však byly právě jen známé korelace. To se změnilo, když se do věci začali míchat fyzikové.
Na druhou stranu víme, že různé vlastnosti organizmů spolu nějak souvisejí. Například větší organizmy se často dožívají vyššího věku, ale mívají méně potomků než ty menší. Takových vzájemných souvislostí známe spoustu, donedávna to však byly právě jen známé korelace. To se změnilo, když se do věci začali míchat fyzikové.
Fyzika a biologie
Vše začalo v polovině devadesátých let 20. století v Santa Fe Institutu v Novém Mexiku. 1) Částicový fyzik Geoff rey West si jednou posteskl, že ho na stará kolena začíná čím dál tím víc zajímat, proč mají různí tvorové tak odlišnou délku života – proč my lidé začínáme rychle stárnout v nějakých sedmdesáti, zatímco želva sloní je v tom věku ještě mladicí. Kolegové jej upozornili, že podobnými otázkami se zabývá o pouhých šedesát mil jižněji, na univerzitě v Albuquerque, ekolog Jim Brown. Ten byl v té době již značně proslulý jako otec makroekologie, tedy oboru zabývajícího se ekologickými procesy a jevy pozorovatelnými na velkých časových a prostorových měřítkách. G. We st s J. Brownem se začali scházet v Santa Fe Institutu, a přestože si zpočátku (podle jejich vlastních slov) vůbec nerozuměli, jejich spolupráce vyústila v jeden z nejgrandióznějších vědeckých projektů posledních desetiletí.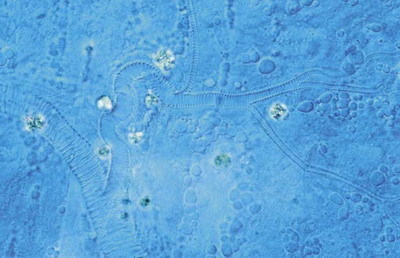 Fyzikální přístup se totiž od biologického značně liší. Zatímco biologové sledují a popisují nejrozmanitější jevy a procesy probíhající v živé přírodě, fyzikové se snaží nacházet obecná, co nejjednodušší a kvantitativně vyjádřitelná pravidla, která by platila nezávisle na kontextu. Tomu v biologii na první pohled brání nesmírná složitost živých systémů. Jenže ve skutečnosti existuje řada fyzikálních systémů, které nejsou méně složité, ale fyzikové je dovedou zjednodušit tak, že vystihnou to podstatné a zanedbají nepodstatné. A o to jde – anebo může jít – i při studiu živých systémů.
Fyzikální přístup se totiž od biologického značně liší. Zatímco biologové sledují a popisují nejrozmanitější jevy a procesy probíhající v živé přírodě, fyzikové se snaží nacházet obecná, co nejjednodušší a kvantitativně vyjádřitelná pravidla, která by platila nezávisle na kontextu. Tomu v biologii na první pohled brání nesmírná složitost živých systémů. Jenže ve skutečnosti existuje řada fyzikálních systémů, které nejsou méně složité, ale fyzikové je dovedou zjednodušit tak, že vystihnou to podstatné a zanedbají nepodstatné. A o to jde – anebo může jít – i při studiu živých systémů.
První, čím se G. West s J. Brownem začali zabývat, byla rychlost metabolizmu. Ta je pro život klíčová, poněvadž – zjednodušeně řečeno – rozhoduje o tom, jak rychle daný organizmus žije, kolik si toho z prostředí bere a kolik toho dává zpět. O rychlosti metabolizmu se vědělo, že těsně souvisí s tělesnou velikostí: větší zvířata mají rychlejší metabolizmus, ale nikoli proporcionálně – vztaženo na jednotku hmotnosti jej mají naopak pomalejší než menší zvířata (viz rámeček 2 ). To se dříve vysvětlovalo poměrem povrchu a objemu: větší zvířata mají relativně menší povrch těla vůči objemu, a tedy teoreticky nižší tepelné ztráty. Tohle vysvětlení by znamenalo, že rychlost metabolizmu bude úměrná hmotnosti umocněné na dvě třetiny (neboť povrch je dvojrozměrný a objem trojrozměrný), ale není to tak: mocnina té funkce se pohybuje někde velmi blízko tří čtvrtin. To znamená, že metabolizmus roste s rostoucí hmotností čím dál tím pomaleji, ale ne zas tak pomalu, aby se to dalo vysvětlit jednoduchou geometrií. K vysvětlení budeme potřebovat o dost složitější geometrii, a hlavně nové pochopení toho, co to organizmus vlastně je.
Co je organizmus
Organizmus se liší od kusu neživé hmoty v několika ohledech. Zaprvé se stará o sebe, tedy dělá to, co je pokud možno prospěšné pro předání svých vlastností do dalších generací. Zadruhé se (přestože tvoří celek) skládá z mnoha částí, z nichž každá v určitém smyslu žije a je důležitá. Každá buňka dělá něco pro celý organizmus, ale o každou buňku se ten organizmus taky musí starat. Jinými slovy je pro organizmus rozhodující neustále budovaný a udržovaný vztah mezi celkem a částmi, a to na mnoha hierarchických úrovních. A teď přijdou značná zjednodušení. Prvním je, že nejdůležitější děje v organizmu probíhají na površích – ať už jde o respiraci, která v posledku probíhá na membránách mitochondrií, nebo o výměnu plynů na povrchu plic či vstřebávání látek v trávicí soustavě. Pokud tedy chceme docílit co největší rychlosti metabolizmu (a to chceme, poněvadž jsme organizmus, který se o sebe stará), chtěli bychom mít také co největší příslušné povrchy, neboť jinak budeme odkázáni na zmíněnou dvoutřetinovou zákonitost pro vztah povrchu a objemu. Můžeme to udělat vcelku jednoduše; nejlíp tak, že zachováme velikost nějakých základních jednotek, na jejichž povrchu příslušné děje probíhají, a velikost těla budeme zvětšovat pouze zvyšováním jejich počtu (což je v podstatě případ buněk). Jejich celkový povrch pak bude růst lineárně s objemem. Jenže to má jednu vadu. Vzhledem k tomu, že organizmus není jen souborem buněk, ale i tím celkem, je třeba nějak zprostředkovat přísun a odsun látek, o něž v metabolizmu jde. Jinými slovy jde nejen o to maximalizovat povrchy, ale také optimalizovat systémy zprostředkující vztah mezi celkem a konečnými povrchy, ale také optimalizovat systémy zprostředkující vztah mezi celkem a konečnými jednotkami metabolizmu, tedy třeba cévní, dýchací nebo vylučovací soustavu. Právě proto, že zprostředkují vztah částí a celku, mají tyto systémy většinou fraktální povahu; jde vlastně o postupně se větvící trubice, u nichž můžeme na každém měřítku pozorovat tentýž průběh větvení (obrázek 1, obrázek 2, obrázek 3, obrázek 5 a obrázek 6).
A teď přijdou značná zjednodušení. Prvním je, že nejdůležitější děje v organizmu probíhají na površích – ať už jde o respiraci, která v posledku probíhá na membránách mitochondrií, nebo o výměnu plynů na povrchu plic či vstřebávání látek v trávicí soustavě. Pokud tedy chceme docílit co největší rychlosti metabolizmu (a to chceme, poněvadž jsme organizmus, který se o sebe stará), chtěli bychom mít také co největší příslušné povrchy, neboť jinak budeme odkázáni na zmíněnou dvoutřetinovou zákonitost pro vztah povrchu a objemu. Můžeme to udělat vcelku jednoduše; nejlíp tak, že zachováme velikost nějakých základních jednotek, na jejichž povrchu příslušné děje probíhají, a velikost těla budeme zvětšovat pouze zvyšováním jejich počtu (což je v podstatě případ buněk). Jejich celkový povrch pak bude růst lineárně s objemem. Jenže to má jednu vadu. Vzhledem k tomu, že organizmus není jen souborem buněk, ale i tím celkem, je třeba nějak zprostředkovat přísun a odsun látek, o něž v metabolizmu jde. Jinými slovy jde nejen o to maximalizovat povrchy, ale také optimalizovat systémy zprostředkující vztah mezi celkem a konečnými povrchy, ale také optimalizovat systémy zprostředkující vztah mezi celkem a konečnými jednotkami metabolizmu, tedy třeba cévní, dýchací nebo vylučovací soustavu. Právě proto, že zprostředkují vztah částí a celku, mají tyto systémy většinou fraktální povahu; jde vlastně o postupně se větvící trubice, u nichž můžeme na každém měřítku pozorovat tentýž průběh větvení (obrázek 1, obrázek 2, obrázek 3, obrázek 5 a obrázek 6).
 Otázka tedy zní: Jak maximalizovat metabolizmus, máme-li k dispozici jednotky konstantní velikosti, jimiž chceme zaplnit většinu objemu organizmu, a přitom potřebujeme minimalizovat ztráty způsobené transportem látek ve fraktálně se větvících systémech? Optimální fraktální rozvodné sítě musí ve svých koncových větvích dosahovat ke všem buňkám, ale přitom musí být co nejkratší a zároveň dobře průchodné (tj. například by se neměly větvit v příliš ostrých úhlech). G. West a J. Brown spolu s botanicky zaměřeným Brianem Enquistem dokázali, že za těchto okolností nelze dosáhnout lepšího výsledku, než je právě ten, pro nějž je charakteristický mocninný vztah mezi hmotností organizmu a rychlostí metabolizmu, kde v oné mocnině jsou tři čtvrtiny. Tohle zjištění publikovali v sérii článků v časopisech Science a Nature, 2) a z nich celá další teorie vychází.
Otázka tedy zní: Jak maximalizovat metabolizmus, máme-li k dispozici jednotky konstantní velikosti, jimiž chceme zaplnit většinu objemu organizmu, a přitom potřebujeme minimalizovat ztráty způsobené transportem látek ve fraktálně se větvících systémech? Optimální fraktální rozvodné sítě musí ve svých koncových větvích dosahovat ke všem buňkám, ale přitom musí být co nejkratší a zároveň dobře průchodné (tj. například by se neměly větvit v příliš ostrých úhlech). G. West a J. Brown spolu s botanicky zaměřeným Brianem Enquistem dokázali, že za těchto okolností nelze dosáhnout lepšího výsledku, než je právě ten, pro nějž je charakteristický mocninný vztah mezi hmotností organizmu a rychlostí metabolizmu, kde v oné mocnině jsou tři čtvrtiny. Tohle zjištění publikovali v sérii článků v časopisech Science a Nature, 2) a z nich celá další teorie vychází.
Dosud jsme mluvili o zvířatech, ale jak je to s rostlinami? Rostliny – alespoň ty větší, jako jsou stromy, keře a také mnohé byliny – jsou v jistém smyslu zvířata naruby. I u nich najdeme fraktální rozvodné systémy, ty ale nejsou uzavřeny dovnitř, nýbrž otevřeny ven. To pochopitelně souvisí s naprosto odlišným způsobem výživy, konkrétně s pohlcováním rozptýlených látek přímo z okolí namísto pohlcování kousků potravy (ostatně různí přisedlí živočichové, kteří se živí podobně, také vypadají spíš jako rostliny). Přitom když si odmyslíme tento rozdíl, jde kupodivu o podobnou situaci jako u živočichů – zase zde jsou nějaké koncové jednotky v principu konstantní velikosti (listy), které je potřeba v prostoru nějak rozmístit a zároveň pro ně najít co nejlepší rozvodné struktury. A opravdu, celý model lze uplatnit i na rostliny, tedy přinejmenším na stromy a keře; u trav nebo pampelišky už by to bylo o dost obtížnější. Horší je, že u rostlin mnohem hůře definujeme jedince.  Naštěstí zde máme obrovskou výhodu, spočívající právě v tom, že rozvodné systémy můžeme uchopit jako fraktály, tedy útvary soběpodobné na různých prostorových měřítkách. Díky tomu se ukazuje, že model funguje, i když třeba místo jednotlivých stromů budeme považovat za jedince jejich jednotlivé větve – ty jsou funkčně zcela rovnocenné stromům příslušné velikosti. A tak se u rostlin nemusíme příliš znepokojovat otázkou, jestli víme, co jsou ti „opravdoví“ jedinci.
Naštěstí zde máme obrovskou výhodu, spočívající právě v tom, že rozvodné systémy můžeme uchopit jako fraktály, tedy útvary soběpodobné na různých prostorových měřítkách. Díky tomu se ukazuje, že model funguje, i když třeba místo jednotlivých stromů budeme považovat za jedince jejich jednotlivé větve – ty jsou funkčně zcela rovnocenné stromům příslušné velikosti. A tak se u rostlin nemusíme příliš znepokojovat otázkou, jestli víme, co jsou ti „opravdoví“ jedinci.
Růst, množení a umírání
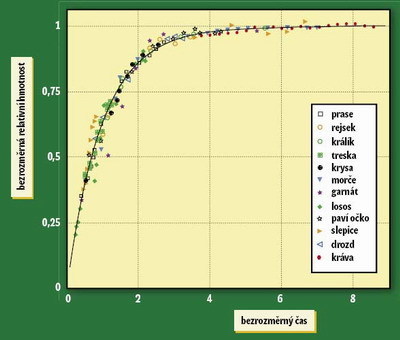
Rychlost metabolizmu podmiňuje rychlost všech biologických procesů. Můžeme proto formulovat předpovědi pro vztah mezi velikostí těla a frekvencí tepu a dýchání (viz rámeček 3 ), ale třeba i rychlostí produkce potomstva (a tedy očekávanou rychlostí populačního růstu). Téže zákonitosti podléhá i délka života. Tím se vyjasňuje prastará záhada, jak je možné, že počet úderů srdce za život zůstává u různých druhů lišících se délkou života přibližně stejný. Můžeme dokonce uvažovat o dvou příčinách, které se navzájem doplňují. První by byla fyziologická (stárnutí organizmu je způsobeno jeho opotřebováním, které je přímo úměrné rychlosti metabolizmu), druhá populačněbiologická: ve víceméně stabilních populacích musí být dlouhodobě vyrovnána porodnost s úmrtností, a pokud produkce potomstva předvídatelně závisí na metabolizmu, potom na něm musí stejně záviset i úmrtnost – jenomže s opačným znaménkem. Menší zvířata (s rychlejší frekvencí tepu) tedy produkují v průměru relativně více potomstva, ale musí o to dříve umírat. To vše jen proto, že na jednotku hmotnosti mají rychlejší metabolizmus. Zajímavou aplikací metabolické teorie je univerzální model individuálního růstu. 3) Každé zvíře roste nejprve rychle, a pak se jeho růst zpomaluje, přičemž některé taxony rostou během života neustále (ryby), jiné ukončí růst velmi brzy a dále se jejich velikost nemění (ptáci). West, Brown a Enquist ukázali, že růst všech živočichů lze charakterizovat jednou jedinou křivkou odvozenou ze základních parametrů buněčného metabolizmu, přičemž organizmy s neukončeným růstem zkrátka nedožijí do doby, kdy se křivka začíná zakřivovat (obrázek). Zákonité zpomalování růstu souvisí právě s tím, že jak organizmus roste, transportní sítě jsou stále složitější, a tím se zpomaluje intenzita metabolizmu vztažená k jednotce hmotnosti.
Zajímavou aplikací metabolické teorie je univerzální model individuálního růstu. 3) Každé zvíře roste nejprve rychle, a pak se jeho růst zpomaluje, přičemž některé taxony rostou během života neustále (ryby), jiné ukončí růst velmi brzy a dále se jejich velikost nemění (ptáci). West, Brown a Enquist ukázali, že růst všech živočichů lze charakterizovat jednou jedinou křivkou odvozenou ze základních parametrů buněčného metabolizmu, přičemž organizmy s neukončeným růstem zkrátka nedožijí do doby, kdy se křivka začíná zakřivovat (obrázek). Zákonité zpomalování růstu souvisí právě s tím, že jak organizmus roste, transportní sítě jsou stále složitější, a tím se zpomaluje intenzita metabolizmu vztažená k jednotce hmotnosti.
Teplota a evoluce
Tělesná velikost není jediným faktorem ovlivňujícím rychlost metabolizmu. Neméně významná je i teplota, poněvadž ta ovlivňuje rychlost všech chemických reakcí, tedy i těch biochemických (viz rámeček 4 ). Vliv teploty na rychlost metabolizmu je opět věc dobře známá, nicméně jeho kvantitativní postižení má zajímavé důsledky. Už se asi nebudeme příliš divit tomu, že teplota předpověditelně ovlivňuje rychlost individuálního vývoje organizmů 4) nebo jejich populačního růstu. 5) Zdá se však, že stejně na teplotě závisí i rychlost evoluce. Přinejmenším molekulární hodiny, charakterizované četností selekčně neutrálních mutací za jednotku času, budou podle teorie tikat rychleji u organizmů žijících v teplejším prostředí (anebo teplokrevných) a taky u těch menších, které mají vyšší rychlost metabolizmu na jednotku hmotnosti. Tím by se mohly rozřešit spory mezi paleontology a molekulárními fylogenetiky týkající se datování evolučních událostí. Molekulární hodiny totiž datují mnohé evoluční události, jako jsou třeba základní rozrůznění ptáků, savců nebo všech mnohobuněčných živočichů, mnohem hlouběji do minulosti, než ukazují fosilní doklady. Možná je to jen tím, že standardně se molekulární hodiny kalibrují na základě fosilně zachycených událostí týkajících se poměrně velkých zvířat, žijících navíc v mírném pásu (poněvadž v tropech, kde žije nejvíc druhů, se fosilie špatně zachovávají). U malých tropických zvířat (kterých je ve skutečnosti většina) mohou molekulární hodiny tikat mnohem rychleji, a tím dávat falešnou představu mnohem delší doby, než která od příslušné evoluční události opravdu uběhla.Z uvedené souvislosti rychlosti evoluce s teplotou je také zřejmé, jak obrovskou výhodu přinesla savcům a ptákům teplokrevnost. Zatímco vnitřní biologický čas studenokrevných tvorů je omezen vnějším prostředím, teplokrevní živočichové jsou z tohoto omezení vyvázáni a jejich vnitřní hodiny tikají poměrně pravidelně a nezávisle na okolí – a navíc vesměs rychleji než u těch studenokrevných. Sami si tak určují svou rychlost evoluce.
Ekosystémy a ekologická sukcese
Ekosystémová ekologie se věnuje koloběhu látek a toku energie v ekosystémech. Po jejím rozkvětu v šedesátých a sedmdesátých letech minulého století nastal jistý úpadek (viz Vesmír 81, 127, 2002/3; 81, 29, 2002/3; 81, 130, 2002/3; 81, 250, 2002/5). Víme sice do nejmenších detailů, jaká je produkce nejrůznějších typů prostředí, jak se na ní podílejí rostliny, živočichové a houby, víme také, jaké množství uhlíku a dusíku kde protéká – jenže nevíme, čím a zda je to vlastně zajímavé. Chybí nám totiž teorie, která by říkala, co máme očekávat, co je banální a co zvláštní. Stejně jako mnohé další vědní obory netrpěla ekosystémová ekologie ani tak nedostatkem empirických zjištění, jako naopak nedostatkem rámce, který by tato zjištění třídil a zařazoval.To se teď možná změní. Víme-li jak spočítat metabolizmus jedince z jeho velikosti a tělesné teploty, mělo by jít spočítat, jaký bude metabolizmus celého ekosystému, pouze na základě klimatických údajů a rozložení populačních početností a tělesných velikostí různých druhů. 6) Jistě, není to tak jednoduché, poněvadž třeba fixace vzdušného uhlíku je jinak citlivá na teplotu než jeho uvolňování do okolí, nicméně pokud o těchto procesech něco víme, dostaneme překvapivě přesné předpovědi třeba koloběhu uhlíku v různých typech prostředí, aniž se potřebujeme dívat na detaily jejich uspořádání. Myšlenka je to jistě zcela jednoduchá, nicméně udělat ten nejjednodušší krok, totiž odvodit metabolizmus celých ekosystémů ze součtu metabolizmu jedinců, kupodivu dosud nikoho nenapadlo.
Podobně je to s vývojem ekosystémů, tedy s ekologickou sukcesí. Už dlouho se ví, že na začátku sukcese převládají rychle rostoucí organizmy s „plýtvavou“ životní strategií, které jsou postupně nahrazovány většími a déležijícími organizmy, přičemž se koloběhy látek a toky energie zpomalují. Bývalo to vysvětlováno téměř mystickým vyzráváním ekosystému až ke konečnému optimálnímu a stabilnímu stadiu nazývanému klimax. Jenže když si uvědomíme, že pro sukcesi je charakteristické hlavně zvětšování průměrné velikosti jedinců spolu s ubýváním jejich počtu, přestaneme se postupnému zpomalování rychlosti metabolizmu celého ekosystému divit. Rychlost sukcese je opět předvídatelná na základě znalosti teploty prostředí a průměrné velikosti jedinců v daném sukcesním stadiu.
Biodiverzita a její trendy
Asi nejpodivnější předpovědi metabolické teorie se týkají rozdílů v počtu druhů mezi prostředími s různou teplotou. Podivné je na tom to, že není žádný zjevný důvod, proč by se prostředí, pro něž je charakteristická různá rychlost metabolizmu (a tedy spotřebovávání a recyklace zdrojů), měla lišit počtem druhů. Druhy se podle metabolické teorie vlastně funkčně moc neliší, nepočítáme-li rozdíly v tělesné velikosti a v trofické úrovni (tedy v tom, jestli jde o rostliny, býložravce či masožravce), a tak by neměl být rozdíl v tom, zda tutéž metabolickou roli hraje jeden nebo sto druhů. Andrew Allen, James Brown a Jamie Gilooly ovšem vycházejí ze zvláštní empirické zákonitosti, z takzvaného pravidla energetické ekvivalence. 7) To říká, že energie, která prochází populací daného druhu na určitém území, nezávisí na jeho tělesné velikosti – oč jsou jedinci menší, o to mají nižší hustoty; sloni toho sežerou víc než myši, ale zase jich není tolik (viz Vesmír 76, 495, 1997/9). Pokud navíc předpokládáme, že by tato energie dále neměla záviset ani na teplotě, znamená to, že v teplejším prostředí musí mít populace jednotlivých druhů nižší hustoty – jinak by každá z nich využívala víc energie než ve studenějším prostředí. Jenže pokud zároveň předpokládáme, že se celkový počet jedinců všech druhů s teplotou nemění, musí být úbytek hustot jednotlivých druhů vyrovnán vyšším počtem druhů.Všechny tyto předpoklady jsou mírně řečeno diskutabilní 8) – proč by se měly organizmy snažit, aby vyhověly pochybnému pravidlu energetické ekvivalence? Proč by v teplejším prostředí nemohly mít jednotlivé druhové populace prostě vyšší celkovou spotřebu energie? Zvláštní ovšem je, že kvantitativně tyto podivné předpovědi překvapivě vycházejí: nejvíc druhů žije v tropech a biodiverzita studenokrevných organizmů roste s teplotou přesně podle této zákonitosti. Autoři pochopitelně vědí, že jejich teorie zatím stojí na velmi vratkých základech. Domnívají se, že za zjištěnými vztahy včetně pravidla energetické ekvivalence stojí spíše zmíněný vztah mezi teplotou a rychlostí evoluce (a tedy i vznikání nových druhů). Ať tak či onak, dobře podložená teorie se zde teprve rodí. Pokračování tohoto příběhu se dozvíme v říjnu v Praze, kde pořádá Santa Fe Institute ve spolupráci s Centrem pro teoretická studia UK a AV ČR pracovní setkání na téma „Scaling biodiversity“, inspirované právě podobnými podivnostmi okolo zákonitostí biologické rozmanitosti.9)
Varování
Jak to tak bývá, hlavní myšlenky metabolické teorie jsou velmi jednoduché. Metabolizmus zkrátka ovlivňuje všechny biologické procesy, a jeho rychlost závisí na tělesné velikosti a teplotě. To už se vědělo. Dnes ale máme k dispozici aparát, díky němuž můžeme rychlost nejrůznějších biologických procesů předpovídat, a to na základě velmi jednoduchých „prvních principů“. 10) Tento aparát přitom není zas tak závislý na základních východiskách celé teorie, tedy na onom fraktálově-optimalizačním pojetí organizmu. Všechny předpovědi by totiž docela dobře fungovaly, i kdyby ony tři čtvrtiny v mocnině vznikaly úplně jinak, třeba dynamikou růstu příslušných transportních systémů (a nešlo by pak vůbec o adaptaci, ale o důsledek ontogenetických procesů). Ostatně podobné tříčtvrtinové zákonitosti najdeme i v neživých systémech, a dokonce i v lidské společnosti (rámeček 5 ). Je tedy dost možné, že jde o něco mnohem univerzálnějšího. To ale nevadí, stejně jako nevadí skutečnost, že ani metabolická teorie zdaleka nevysvětlí veškerou proměnlivost projevů života. Důležité je, že máme teo rii, která produkuje testovatelné hypotézy, teorii, od níž lze hledat odchylky a kterou lze vyvracet. Takové pokusy se vskutku dějí 11) a čas ukáže, nakolik byly úspěšné.Metabolická teorie tedy umožňuje vidět fenomény, které by jinak zůstaly skryté, umožňuje kvantitativně předpovídat jevy od molekulárních dějů v buňce až po dynamiku celých ekosystémů – a pak se třeba divit, že ty předpovědi nesedí. Je ale výjimečná také tím, že do centra svého zájmu staví nikoli molekuly nebo buňky, ale jedince. Jedinci se stávají znovu centrálními jednotkami biologických procesů, a nezáleží na tom, že mnohdy přesně nevíme, co to vlastně jedinec je.
Poznámky
SLOVNÍČEK
Boltzmannova konstanta – konstanta vztahující změny teploty k změnám energie molekul
mitochondrie – buněčné organely uplatňující se v energetickém metabolizmu
respirace – dýchání; buněčný proces, při němž je získávána energie, spotřebováván kyslík a produkován oxid uhličitý
selekčně neutrální mutace – takové, které jsou zanedbatelné z hlediska přirozeného výběru
sukcese – vývoj společenstva, spočívající v postupném a jednosměrném nahrazování populacemi určitých druhů populacemi jiných druhů
taxon – skupina příbuzných druhů pocházejících ze společného předka
BIOLOGIE VZTAH METABOLIZMU A TĚLESNÉ HMOTNOSTI
Když vyneseme vztah mezi intenzitou metabolizmu a tělesnou hmotností v logaritmickém měřítku, dostaneme přímku se sklonem 0,75.
Rovnice této přímky je
lnP = 0,75 . lnM + lnM0 ,
kde P je intenzita metabolizmu (jde vlastně o energetický příkon, proto P jako „power“), M je hmotnost jedince a M0 je konstanta specifi cká pro daný taxon (například savce). Po odlogaritmování dostáváme mocninnou funkci P = M0 . M0,75 .
Sklon přímky v logaritmicko-logaritmickém měřítku se zde tedy stává mocninou. Jde o bezrozměrné číslo (v tomto případě 0,75), které nám může napovědět, co je za pozorovaný jev odpovědné. Protože sklon přímky (a tedy exponent v závislosti intenzity metabolizmu na hmotnosti) je menší než 1, dochází s růstem hmotnosti ke „zpomalení“ intenzity metabolizmu.
Mocninné funkce obecně poukazují k měřítkové invarianci, tedy k soběpodobnosti a fraktálům v nejširším slova smyslu. Platí pro ně totiž – laicky řečeno – že nárůst veličiny na ose x na nějaký násobek původní hodnoty vede k nárůstu příslušné funkční hodnoty (na ose y) o její konstantní násobek, nezávisle na tom, jaké jsou absolutní velikosti příslušných veličin. To právěvyjadřuje přímka v logaritmicko-logaritmickém měřítku.
METABOLIZMUS A OSTATNÍ RYCHLOSTI
Víme-li, že P ~ M3/4, rychlost metabolizmu na jednotku hmotnosti B bude s hmotností klesat podle vztahu B ~ M–1/4, poněvadž B = P/M, a tedy B ~ M3/4 .M–1 = M–1/4. Totéž se pak bude týkat frekvence biologických dějů, neboť ty jsou přímo úměrné rychlosti metabolizmu. Naopak jejich trvání (třeba doba obměny metabolických substrátů, a vposledku i doba života) bude růst s hmotností tak, že t ~ M1/4, protože jde o převrácenou hodnotu frekvence
TEPLOTA A BIOCHEMICKÁ KINETIKA
Biochemickou kinetiku popisuje exponenciální Van’t Hoff ova – Arrheniova rovnice
B ~ e–E/kT,
kde k je Boltzmannova konstanta, T je teplota v kelvinech a E je průměrná aktivační teplota metabolizmu (o níž bylo zjištěno, že zůstává víceméně konstantní pro všechny organizmy, což evidentně souvisí s tím, že většina základních biochemických dějů je společná). Vztah vychází ze skutečnosti, že s rostoucí teplotou roste podíl molekul, jejichž kinetická energie je již dostatečná k tomu, aby vstoupily do příslušných chemických reakcí. Kombinaci vlivu velikosti i teploty na metabolizmus jedince lze tedy postihnout uni- verzální rovnicí ve tvaru
P = P0 . M3/4 . e–E/kT,
kde P0 je konstanta specifi cká pro daný taxon.
LIDSKÉ SPOLEČNOSTI, ENERGIE A PORODNOST
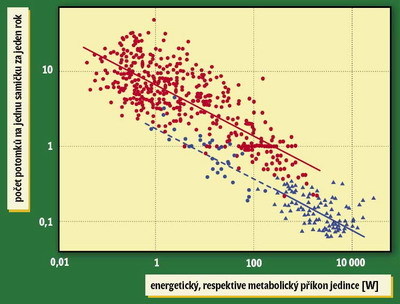 Pokud předpokládáme, že energetický příkon P ~ M3/4 a natalita (produkce potomstva na jedince) R souvisí s hmotností tak, že R ~ M–1/4, můžeme oba vztahy zkombinovat a získat vztah mezi příkonem a porodností R ~ P–1/3 .
Pokud předpokládáme, že energetický příkon P ~ M3/4 a natalita (produkce potomstva na jedince) R souvisí s hmotností tak, že R ~ M–1/4, můžeme oba vztahy zkombinovat a získat vztah mezi příkonem a porodností R ~ P–1/3 .
Tento vztah například v rámci savců opravdu platí; čím je savec větší a má vyšší spotřebu energie, tím mívá méně potomků. Naprosto šokující ale je, že pro lidské společnosti platí úplně tentýž vztah: čím jsou bohatší, tím mají nižší porodnost (viz Vesmír 83, 130, 2004/3), a když do jednoho grafu vyneseme jednotlivé druhy savců (kolečka) i jednotlivé lidské společnosti (trojúhelníčky), většina bodů bude ležet na jedné přímce. Přesně řečeno, lidské společnosti budou ležet na přímce platné pro primáty (modrá kolečka), která je trochu posunutá dolů vůči přímce platné pro všechny savce.
Tentýž sklon získáme, dokonce i když si proti sobě vyneseme spotřebu energie a porodnost v jediné zemi, ale v různých letech. Jediné, čím se lidské společnosti od savčích druhů liší, je to, že jejich spotřeba je mnohem vyšší (a porodnost mnohem nižší), než by odpovídalo savci příslušné velikosti – člověk z vyspělé společnosti by svou spotřebou a porodností odpovídal třicetitunovému primátovi. Zatímco prostý metabolický příkon člověka je pouhých 100 W (tedy jako žárovka), jeho spotřeba energie je díky civilizačním vymoženostem stokrát vyšší (Moses M. E., Brown J. H., Ecology Letters 6, 295–300, 2003).
Proč platí pro lidské společnosti tentýž vztah jako pro biologické druhy, zůstává záhadou. Nemůže jít o stejné vysvětlení jako v případě metabolizmu jedinců, poněvadž v lidských společnostech se většina spotřeby energie uskutečňuje provozem nejrůznějších strojů a dalších zařízení. Autoři spekulují o tom, že by „rozvodné sítě“ v lidských společnostech (dopravní tepny, elektrorozvodné sítě) mohly podléhat těm samým zákonitostem jako transportní systémy v těle, nicméně inženýrské sítě většinou nemají tak úplně fraktální povahu, a hlavně není jasné, jak a proč by se to mělo nakonec odrážet v porodnosti. Zdá se, že za vším bude ještě nějaká hlubší, zatím nepoznaná zákonitost.
Ke stažení
 článek v pdf souboru [1,36 MB]
článek v pdf souboru [1,36 MB]
O autorovi
David Storch

Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
