









Promiskuitní DNA
Genetická informace je v eukaryotických buňkách uložena nejen v jádře. Svou vlastní DNA obsahují také malé buněčné organely – mitochondrie a chloroplasty. Podle endosymbiotické teorie se tyto organely kdysi vyvinuly z volně žijících bakterií a z doby svého vzniku si uchovaly zbytky genetické informace (viz Markoš A.: Třicet let endosymbiotické teorie, Vesmír 78, 208, 1999/4).
Pozůstatky prokaryotických genomů
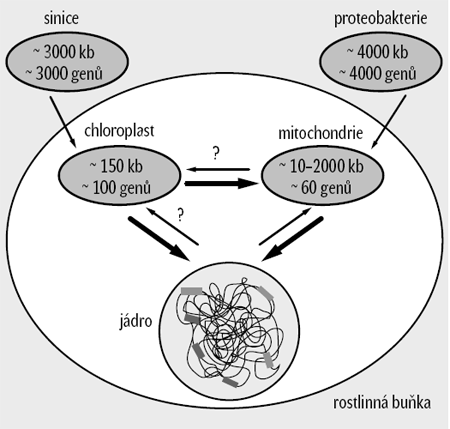 Primitivní buňka, buď prokaryotická, nebo již eukaryotická, 1) přijala nejprve bakterie, 2) a z nich se vyvinuly mitochondrie. Poté tato buňka, tehdy zřejmě bičíkatá, přijala další bakterie, 3) které daly vzniknout chloroplastům. Tyto organismy stály u zrodu říše rostlin. Během evoluce se organelové genomy drasticky zmenšily. Zatímco prokaryotičtí předchůdci organel měli genomy o milionech bází, genomy organel jsou o jeden až dva řády menší (obrázek 1). Průměrný chloroplastový genom má velikost kolem 150 kilobází, mitochondriální genomy živočichů mívají desítky kilobází (ten lidský má 16,6 kilobáze a obsahuje 37 genů), rostlinné mitochondriální genomy jsou podstatně větší a dosahují stovek až tisíců kilobází. Spolu s redukcí velikosti genomů klesl počet genů v genomech organel, z původních několika tisíc genů kódujících proteiny jich zbylo několik desítek, nanejvýš několik set. Některé linie chloroplastů ztratily více než 95 % genů, tj. sekvencí kódujících ve volně žijících sinicích proteiny a různé typy RNA. Kam se ztracené geny poděly?
Primitivní buňka, buď prokaryotická, nebo již eukaryotická, 1) přijala nejprve bakterie, 2) a z nich se vyvinuly mitochondrie. Poté tato buňka, tehdy zřejmě bičíkatá, přijala další bakterie, 3) které daly vzniknout chloroplastům. Tyto organismy stály u zrodu říše rostlin. Během evoluce se organelové genomy drasticky zmenšily. Zatímco prokaryotičtí předchůdci organel měli genomy o milionech bází, genomy organel jsou o jeden až dva řády menší (obrázek 1). Průměrný chloroplastový genom má velikost kolem 150 kilobází, mitochondriální genomy živočichů mívají desítky kilobází (ten lidský má 16,6 kilobáze a obsahuje 37 genů), rostlinné mitochondriální genomy jsou podstatně větší a dosahují stovek až tisíců kilobází. Spolu s redukcí velikosti genomů klesl počet genů v genomech organel, z původních několika tisíc genů kódujících proteiny jich zbylo několik desítek, nanejvýš několik set. Některé linie chloroplastů ztratily více než 95 % genů, tj. sekvencí kódujících ve volně žijících sinicích proteiny a různé typy RNA. Kam se ztracené geny poděly?
Objev promiskuitní DNA
Dříve se badatelé domnívali, že genetické systémy eukaryontních buněk – dva u živočichů (jádro a mitochondrie) a tři u rostlin (jádro, mitochondrie a chloroplasty) – jsou oddělené a každý si jde svou evoluční cestou. Na počátku osmdesátých let však vědci objevili chloroplastové geny v mitochondriálním genomu. Bylo jasné, že se tam musely dostat přenosem genů mezi organelami. Některé úseky chloroplastové DNA byly dokonce v mitochondriálním genomu obsaženy vícekrát, což svědčilo o vícenásobném přenosu. Tehdy vznikl termín „promiskuitní DNA“, označující přenos DNA mezi buněčnými genomy. Objev promiskuitní DNA vyvolal řadu otázek – zda se také mitochondriální geny mohou nacházet v chloroplastech, jak daleko tato promiskuita DNA vůbec může jít, zda něco podobného může nastat i u vyšších organismů, například u člověka a jeho symbiotických organismů, či dokonce u nesymbiotických patogenních organismů. Výsledky sekvencování celých genomů dnes již jasně ukazují, že přenos genů organel do jádra je všudypřítomný a vyskytuje se častěji, než jsme si dříve dokázali představit.Proč se geny přesouvají do jádra?
Prokaryotičtí předchůdci organel byli původně sexuální, a poté, co vstoupili do buňky a uzavřeli endosymbiotické partnerství, se od ostatních jedinců svého druhu izolovali, začali být asexuální. Víme, že nepřítomnost sexuality vede k hromadění škodlivých mutací. Nefungují totiž opravné mechanismy na bázi rekombinace DNA, které spočívají v opravě poškozených kopií genů podle kopií nepoškozených. Selekční tlak kromě toho zvýhodňuje malé genomy. V evoluci byly zvýhodněny úseky DNA, které byly přeneseny do jádra, kde mohou rekombinovat, a tím opravovat své poškození. Ukázalo se, že během evoluce organel rostlinných buněk a řas (plastidů) bylo do jádra přeneseno až 5000 genů. Mnohé kopie těchto genů jsou funkční i po přenosu do jádra. Organely totiž využívají produkty svých bývalých genů, i když jsou tyto geny v jádře.Které geny jsou promiskuitnější?
Proč některé geny v organelách zůstávají? Byly totiž nalezeny i organely prokazatelně endosymbiotického původu, které ztratily DNA. Odpověď zatím není jednoznačná, ale zdá se, že některé geny zůstaly v organelách zachovány v důsledku určitých vlastností proteinů (hydrofobicitou), a také v souvislosti s řízením oxidativních procesů. Některé proteiny by těžko procházely membránami z cytoplazmy do organel, a proto je výhodnější je syntetizovat přímo uvnitř organel. Stejně tak v případě proteinů řídících oxidativní procesy je výhodnější jejich syntéza přímo v organelách, kde mohou být odpovídající geny zapínány a vypínány podle potřeby organel. Ukázalo se, že v evoluci se některé geny z organel přenášejí do jádra přednostně. Co je příčinou různé náchylnosti genů k promiskuitě? Jako první přecházejí do jádra geny, jejichž produkty mají regulační funkci. Geny související převážně s překladem genetické informace a s dýcháním opouštějí organely jako poslední. Pěkným příkladem je enzym rubisco, jehož katalytická podjednotka je kódována chloroplastem, zatímco regulační podjednotka pochází z jádra (obrázek 2).Jak se organelové geny dostanou do jádra?
Původně se uvažovalo o prostřednictví genetických elementů, jako jsou transpozony, nebo o fágovém přenosu. Dnes existenci organelové DNA v jádře vysvětlují dvě hypotézy: přenos prostřednictvím RNA a včlenění velkých kusů organelové DNA rekombinací. Tyto hypotézy se navzájem nevylučují a pro obě existují experimentální důkazy. V čem spočívá první hypotéza? Organelový gen se přepíše do RNA, a pak se zpětně přepíše (reverzní transkriptázou) do komplementární DNA (cDNA), která se včlení do jaderného genomu. Skutečně byly nalezeny úseky organelových genů v jádře, jimž chyběly introny, tj. části genů, které zmizí pouze při úpravách RNA. Představu včleňování velkých kusů DNA podpořily objevy 131 kilobází úseku chloroplastové DNA v jádře rýže nebo 620 kilobází úseku mitochondriální DNA v jádře huseníčku (Arabidopsis thaliana). Úseky obsahovaly nejen geny s introny, ale i mezigenové oblasti. Tyto obří kusy DNA byly zřejmě procesem rekombinace včleněny do jaderného genomu poté, co se membrána organel rozpustila a jejich DNA se dostala do cytoplazmy a do jádra.Jak častý je přenos genů do jádra?
 Vědci se rozhodli, že to zjistí. Vložili proto do chloroplastového genomu tabáku gen kódující rezistenci k antibiotiku a zajistili, aby byl tento gen zapnut jen tehdy, když je v jádře. Poté prokázali, že u rostlin tabáku, jež vyrostly v přítomnosti daného antibiotika, se do jádra přenesl kousek chloroplastového genomu nesoucího rezistenci k antibiotiku. S překvapením zjistili, že je takový přenos poměrně častý, u jedné ze 16 000 rostlin. Ve skutečnosti je ale mnohem častější, neboť výše uvedeným postupem byla sledována pouze část chloroplastového genomu, která obsahovala gen pro rezistenci. Lze říci, že z několika tisíc rostlin, které vidíme, když se kolem sebe rozhlédneme například na palouku v lese, nese alespoň jedna v jádře „čerstvý“ kousek chloroplastové DNA, který se do jádra včlenil v minulé generaci.
Vědci se rozhodli, že to zjistí. Vložili proto do chloroplastového genomu tabáku gen kódující rezistenci k antibiotiku a zajistili, aby byl tento gen zapnut jen tehdy, když je v jádře. Poté prokázali, že u rostlin tabáku, jež vyrostly v přítomnosti daného antibiotika, se do jádra přenesl kousek chloroplastového genomu nesoucího rezistenci k antibiotiku. S překvapením zjistili, že je takový přenos poměrně častý, u jedné ze 16 000 rostlin. Ve skutečnosti je ale mnohem častější, neboť výše uvedeným postupem byla sledována pouze část chloroplastového genomu, která obsahovala gen pro rezistenci. Lze říci, že z několika tisíc rostlin, které vidíme, když se kolem sebe rozhlédneme například na palouku v lese, nese alespoň jedna v jádře „čerstvý“ kousek chloroplastové DNA, který se do jádra včlenil v minulé generaci.
Promiskuitní DNA v lidském genomu
Úseky promiskuitní DNA jsou i v jaderném genomu člověka. Kromě mitochondriálních genů, které se přesunuly do jádra před několika stovkami milionů let a konají svoji práci pod novou vládou, bylo v našem genomu identifikováno přes 200 nefunkčních fragmentů mitochondriální DNA. Z nich vědci dále zkoumali 42 úseků, které se do našeho jaderného genomu včlenily nejpozději. Zjistili, že pouze 14 z nich se nachází také v jaderném genomu šimpanze, což naznačuje, že většina těchto úseků byla do našeho genomu včleněna až po oddělení větví vedoucích k člověku a k šimpanzům, tedy někdy před 4–8 miliony let. Některé úseky se dokonce liší i mezi lidskými populacemi, to znamená, že byly včleněny „nedávno“. Fragmenty mitochondriální DNA nejsou v lidském genomu rozptýleny rovnoměrně. Pohlavní chromozom Y, přítomný pouze u mužů, je mitochondriální DNA kolonizován mnohem více než ostatní chromozomy. Zajímavé je i to, že se promiskuitní DNA včleňuje s mnohem vyšší frekvencí do vysoce aktivních genů než do oblastí nekódujících (např. repetitivních) sekvencí. Nejnovější objevy naznačují, že úseky této promiskuitní DNA v lidském genomu nejsou zcela neškodné. Jejich včlenění do genů může způsobit i závažná onemocnění, což dokazuje nedávný objev mitochondriální DNA v genu důležitém pro srážlivost krve u pacientů trpících hemofilií.Promiskuitní DNA všude
Organelová DNA se pohybuje z efektivně využitého genomu prokaryot do zcela odlišného genetického prostředí eukaryotického jádra, obsahujícího složené geny a velké množství opakujících se úseků DNA, repeticí. Organely se vyznačují nejen odlišnou genovou i genomovou organizací ve srovnání s jádrem, ale i odlišnými mechanismy regulace genové exprese. Podíl promiskuitní DNA v genomech byl dosud podhodnocován, což bylo způsobeno tím, že algoritmy používané v sekvenačních projektech považovaly organelové sekvence za nežádoucí kontaminace a systematicky je vyřazovaly. To se nyní přehodnocuje. Ukázalo se, že například v genomu huseníčku (Arabidopsis thaliana) pochází 18 % genů ze starobylého plastidového genomu. Organismy se brání, aby jejich jaderné genomy nebyly přesyceny organelovými sekvencemi. Aktivně se jich zbavují různými mechanismy, především rekombinací. V laboratoři vývojové genetiky rostlin v Biofyzikálním ústavu AV ČR, kde se zabýváme strukturou a evolucí pohlavních chromozomů u rostlin, jsme zjistili, že se na pohlavním chromozomu Y modelové dvoudomé rostliny silenky širolisté (Silene latifolia) hromadí chloroplastová DNA. Zřejmě je to tím, že u chromozomu Y, který nemá partnera (na rozdíl od autozomů a pohlavního chromozomu X), a tudíž nerekombinuje, tento obranný mechanismus nefunguje. Ukazuje se, že všudypřítomný přenos genů z organel do jádra a vzájemné ovlivňování mezi buněčnými genomy hrají v evoluci genomů důležitou úlohu. Spolu s přenosnými elementy – především s retroelementy (viz Vesmír 79, 273, 2000/5) – je jedním z hlavních činitelů ovlivňujících dynamiku eukaryontních genomů od jejich vzniku.Poznámky
SLOVNÍČEK
autozom – nepohlavní chromozom vyskytující se v párech v somatických buňkách. Autozomy jsou stejné u obou pohlaví.endosymbiotická teorie vzniku eukaryotické buňky – starobylé buňky, které byly podobné dnešním archebakteriím, v minulosti pohltily jiné bakterie, a posléze si je „ochočily“. Z pohlcených bakterií pak vznikly buněčné organely: z proteobakterií mitochondrie (v živočišných i rostlinných buňkách), ze sinic chloroplasty (jen v rostlinných buňkách).
plastidy – organely rostlinných buněk a řas. Většinou se předpokládá, že jde o potomky bývalých endosymbiontů sinicového typu, v několika málo případech u konkrétních druhů řas však existuje podezření, že jejich plastidy jsou potomky eukaryotických symbiontů typu řas.
V buňce slouží plastid jako organela, v níž probíhá fotosyntéza, jako zásobní organela, nebo jako organela odpovědná za zbarvení buňky. DNA obsahuje některé geny, plastidy tedy mají důležitý podíl na mimojaderné dědičnosti. Mohou nabývat různých forem v závislosti na funkci, kterou v rostlině plní. Některé formy nejsou definitivní a mohou přecházet v jiné.
promiskuitní DNA – úseky DNA migrující v průběhu evoluce mezi genomy buněčných organel (mitochondrií a chloroplastů) a jaderným genomem.
rekombinace – vytváření nových kombinací genů či jiných sekvencí DNA, jehož základem je překřížení odpovídajících částí chromozomů (crossing-over), při němž se molekuly DNA „zlomí“ a opět se spojí s novými kombinacemi. Rekombinující oblasti jsou většinou homologické, není to však podmínkou. Mechanizmus rekombinace se uplatňuje zejména při výměně genetického materiálu mezi rodičovskými chromozomy při tvorbě pohlavních buněk v meióze.
repetitivní sekvence – mnohonásobně opakovaná sekvence nukleotidů v DNA. Takové sekvence představují značnou část lidského genomu.
retroelementy – genetické elementy, které k své produkci používají enzym reverzní transkriptázu a kopírují se prostřednictvím molekul RNA (podrobněji viz E. Kejnovský: Zkopíruj a ulož, Vesmír 79, 273, 2000/5).
reverzní transkriptáza – enzym tvořící molekulu DNA na základě přepisu informace uložené v RNA (opačně než při běžném přepisu).
složené geny – geny, jejichž části (tvořící dohromady většinu délky genu) se po přepisu z molekuly DNA do RNA odstraní ještě před vznikem proteinu. Taková je většina genů eukaryotických organizmů.
Ke stažení
 článek ve formátu pdf [510,31 kB]
článek ve formátu pdf [510,31 kB]
O autorovi
Eduard Kejnovský

Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
