









Myš domácí
 Když deset let po výstřelu z Aurory dva ruští emigranti zveřejnili ve Zprávách Francouzské akademie věd svůj článek o rozmnožování bezocasých myší, netušili, že náhodou zakopli o fascinující „organizmus v organizmu“, který je dnes jedním z nejznámějších a nejlépe prozkoumaných „ultrasobeckých“ genetických elementů.
Když deset let po výstřelu z Aurory dva ruští emigranti zveřejnili ve Zprávách Francouzské akademie věd svůj článek o rozmnožování bezocasých myší, netušili, že náhodou zakopli o fascinující „organizmus v organizmu“, který je dnes jedním z nejznámějších a nejlépe prozkoumaných „ultrasobeckých“ genetických elementů.
Některé myši na jednom ze svých chromozomů nesou komplex genů, které se lidově řečeno „udělaly samy pro sebe“ a začaly se sobecky šířit na úkor genů ostatních. 2) Ačkoli jsou tito „odpadlíci“ desetkrát větší než celá DNA bakterie E. coli, až na výjimky jsou „neviditelní“, neboť jejich nositele nelze vystopovat podle zjevných morfologických odchylek. Jsou v genomu domácí myši něčím, co lze přirovnat k trojskému koni plnému genetických vojáků, kteří číhají na příležitost pobít své konkurenty.
S jistým zjednodušením lze počátek historie výzkumu této struktury vystopovat v Paříži mezi dvěma světovými válkami, kde r. 1927 ruští emigranti N. Dobrovolskaja-Zavadskaja a N. Kobozieff publikovali výsledky svého výzkumu bezocasých variant myší pod titulkem Sur la reproduction des Souris anoures (O reprodukci bezocasých myší). Aniž to tušili, setkali se s fenoménem, který se jako červená nit vine souborem nejrozmanitějších fenotypových projevů – od účinků na embryonální vývoj, plodnost samců a genetickou rekombinaci až po narušení druhého Mendelova zákona, strukturu nazývanou t-haplotyp (nebo ve starší literatuře t-mutace či t-alela).
Ještě poměrně nedávno byly t-haplotypy pro biology záhadou. Široké spektrum jejich účinků vyvolávalo řadu otázek: Co je podstatou t-haplotypů? Které okolnosti je vyvolávají a jaké mechanizmy ovlivňují tolik vzájemně nepříbuzných vlastností jejich nositelů? Jak t-haplotypy vznikají a jak se udržují v přírodních populacích?
-haplotyp je když…
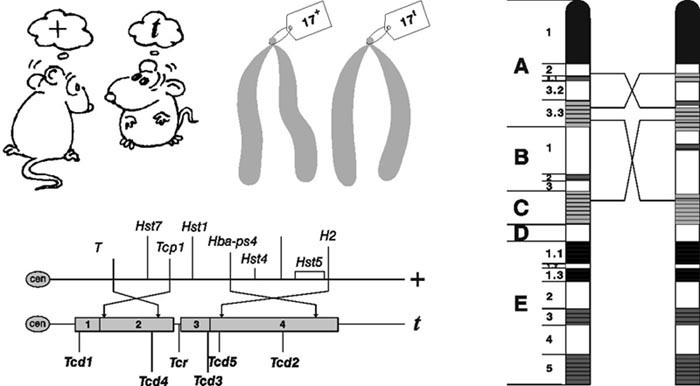
Co vlastně máme na mysli, jestliže hovoříme o t-haplotypu? Ačkoli již před více než třiceti lety publikovala britská genetička Mary Lyonová v sérii článků názor, že t-haplotypy ve skutečnosti představují větší genetickou jednotku s pozměněnou chromozomální strukturou, kolegové její práce ignorovali a všechny mutantní fenotypové projevy připisovali mnohačetným účinkům jediného genu. Teprve se zavedením přesnějších cytogenetických a molekulárněgenetických metod byla odhalena detailní struktura celého komplexu a Lyonová se dočkala zadostiučinění. (V této souvislosti připomeňme osudy dalších žen ve vědě, např. Barbary McClintockové, objevitelky „skákajících genů“ čili transpozonů, nebo Lynn Margulisové, jejíž jméno je spojeno s endosymbiotickou teorií původu buněčných organel.)Celý t-komplex zaujímá oblast asi 20 centimorganů 5) v proximální (bližší k centromeře) části chromozomu 17 a odpovídá asi 30–40 milionům bází (pro srovnání: mitochondriální gen pro cytochrom b má přibližně 1000 bází). Jeho délka představuje zhruba jednu třetinu chromozomu 17, tedy 1,2 % veškeré myší jaderné DNA. Za svůj název vděčí t-haplotyp přítomnosti genu T (tailess) odpovědného za myší „krátkoocasost“ (brachyurii). Ve skutečnosti je celá situace poněkud složitější – za zmiňovaný fenotypový projev může nulová mutace tohoto genu na jinak normálním chromozomu 17, která příslušný gen „vypne“. Zatímco kombinace této mutace s nemutovaným „kolegou“ (T/+) 3) vede k vývoji krátkého ocasu, jedinci s jednou kopií t-haplotypu (T/t) nemají ocas žádný.
 Termín „haplotyp“ označuje skutečnost, že se celý komplex, obsahující více než 560 genů, chová jako „jeden muž“. Mají to na svědomí čtyři chromozomové přestavby – paracentrické inverze. (Představme si vyrovnanou řadu vojáků, z níž postupně čtyři muži postoupí o krok vpřed, udělají čelem vzad a znovu se zařadí, obráceni o 180° oproti svým spolubojovníkům. Inverze se ovšem týkají různě dlouhých úseků chromozomů, obsahujících většinou desítky genů.) Jak nás poučí každá učebnice genetiky, tento typ mutace vyvolává při párování homologických chromozomů v průběhu meiózy (redukčního buněčného dělení, při němž vznikají zárodečné buňky) inverzní smyčku. Ta zabraňuje crossing-overu (výměně nukleotidových sekvencí) a rekombinaci, což ve svém důsledku brání „rozbití“ celého komplexu a ztrátě příslušných vlastností. Čtyři po sobě jdoucí inverze přemění celý komplex v poměrně kompaktní monolitickou strukturu.
Termín „haplotyp“ označuje skutečnost, že se celý komplex, obsahující více než 560 genů, chová jako „jeden muž“. Mají to na svědomí čtyři chromozomové přestavby – paracentrické inverze. (Představme si vyrovnanou řadu vojáků, z níž postupně čtyři muži postoupí o krok vpřed, udělají čelem vzad a znovu se zařadí, obráceni o 180° oproti svým spolubojovníkům. Inverze se ovšem týkají různě dlouhých úseků chromozomů, obsahujících většinou desítky genů.) Jak nás poučí každá učebnice genetiky, tento typ mutace vyvolává při párování homologických chromozomů v průběhu meiózy (redukčního buněčného dělení, při němž vznikají zárodečné buňky) inverzní smyčku. Ta zabraňuje crossing-overu (výměně nukleotidových sekvencí) a rekombinaci, což ve svém důsledku brání „rozbití“ celého komplexu a ztrátě příslušných vlastností. Čtyři po sobě jdoucí inverze přemění celý komplex v poměrně kompaktní monolitickou strukturu.
-haplotyp působí?
Působení t-haplotypů lze shrnout do tří kategorií. Zatímco první dva důsledky – sterilita a embryonální úmrtnost (letalita) – se projevují v životních funkcích celého organizmu, třetí z nich, z evolučního hlediska nejzajímavější, je mnohem subtilnější. Nazývá se „vychýlení transmisního poměru“ (transmission ratio distortion). Oč jde? Jak říká první Mendelův zákon, jednotlivé vlohy volně segregují, což převedeno do lidského jazyka znamená, že do každé pohlavní buňky (ta je haploidní – má jen jednu sadu chromozomů) vstoupí znak zděděný buď od matky, nebo od otce, a to se stejnou pravděpodobností. V našem případě tedy zhruba polovina pohlavních buněk samce t/+ ponese chromozom 17 „divokého typu“, zatímco druhá polovina se bude pyšnit mutantním chromozomem obsahujícím t-haplotyp. Dalo by se logicky očekávat, že pokud tento samec zplodí potomstvo se samicí, která má stejnou genetickou dispozici (tj. t/+), bude výskyt t-haplotypu opět (plus minus) padesátiprocentní. Ale chyba lávky! Ve skutečnosti má t-haplotyp v buňkách mnohem více potomků, než velí teoretický předpoklad – udává se, že transmisní poměr může být vychýlen až přes 95 % ve prospěch alely t.Tento jev není v přírodě ojedinělý. Mezi nejznámější příbuzné myšího t-haplotypu patří geny nazývané souhrnně segregation distorters (SD) u octomilky Drosophila melanogaster (viz Vesmír 77, 67, 1998/2) a spore killers u houby Neurospora crassa. Avšak ačkoli funkčně jsou tyto systémy analogické, každý z nich působí na základě odlišných molekulárních mechanizmů. Obecně se takový přenos genetických elementů do dalších generací označuje jako meiotický tah 4) a může se týkat jednotlivých genů, skupin genů či celých chromozomů (viz např. robertsonské chromozomy u rejska obecného a pravděpodobně i u myši domácí či přídatné B-chromozomy u myšic rodu Apodemus). Časté jsou rovněž případy podobného nelítostného soupeření mezi samčími a samičími geny, které vedou k vychýlení poměru pohlaví. Případy, kdy se určitý genetický element odmítne řídit zavedenými, všeobecně respektovanými pravidly a začne se chovat na úkor ostatních, rozhodně nejsou nahodilou hříčkou přírody poskytující profesionální potěchu několika kabinetním učencům nebo vyvolávající lehké mrazení v zádech čtenářů sobotních novinových příloh. Naopak lze očekávat, že s postupným zaváděním přesnějších metod a rozšířením spektra zkoumaných organizmů bude počet prokázaných případů takového přenosu částí genomu narůstat.
 Vraťme se ale k domácím myším a t-haplotypu. Ještě než si alespoň v hrubých rysech načrtneme předpokládaný molekulární mechanizmus vychýlení transmisního poměru, je třeba se na chvíli zastavit u pojmu „meiotický“ – ten v tomto případě platí jen zčásti. I když je celý proces (týkající se v tomto případě pouze samců) nastartován skutečně během meiotického párování homologických chromozomů, vlastní účinek se projeví až během diferenciace samčích pohlavních buněk a při závodu o možnost splynutí se samičím vajíčkem.
Vraťme se ale k domácím myším a t-haplotypu. Ještě než si alespoň v hrubých rysech načrtneme předpokládaný molekulární mechanizmus vychýlení transmisního poměru, je třeba se na chvíli zastavit u pojmu „meiotický“ – ten v tomto případě platí jen zčásti. I když je celý proces (týkající se v tomto případě pouze samců) nastartován skutečně během meiotického párování homologických chromozomů, vlastní účinek se projeví až během diferenciace samčích pohlavních buněk a při závodu o možnost splynutí se samičím vajíčkem.
Dosud bylo nalezeno celkem šest genů odpovědných za vychýlení transmisního poměru. Pět z nich, označovaných zkratkou Tcd (t complex distorter), je obdobou systému genů SD u octomilky. Tím šestým je gen Tcr (t complex responder), který podobně jako u octomilky hraje v celém procesu klíčovou roli (na obrázku). Původní divoká forma toho genu, kterou si můžeme pro přehlednost označit jako R+, je citlivá vůči působení genů Tcd. Jejich účinek se projeví poruchami procesu zrání spermií a narušením jejich schopnosti účastnit se oplození vajíčka. Výsledkem je skutečnost, že drtivá většina potomků nese kopii t-haplotypu. Těžko si lze představit důmyslnější, a hlavně účinnější mechanizmus vlastního přežití! (Asi znáte rčení, že „nejlepší obranou je útok“.) Mutantní forma klíčového šestého genu, řekněme Rt, je vůči vlivům zbývajících pěti genů – distorterů (deformátorů, narušitelů) – poměrně odolná. Odolnost však není absolutní, takže v případě, že jsou tyto geny v homozygotním stavu (sejdou se během meiózy dva chromozomy nesoucí t-haplotyp), budou jejich nositelé reprodukčně sterilní. S trochou nadsázky si to můžeme představit tak, že zrající spermie nesoucí thaplotyp svého meiotického partnera „usmrtí“ tak jako tak: u heterozygotního samce jsou eliminovány chromozomy divokého typu, zatímco u samce se dvěma kompletními t-haplotypy se pohlavní buňky „povraždí“ navzájem a nevyhnutelně tak způsobí jeho neplodnost.
Mechanizmus je ve skutečnosti složitější, např. geny Tcd se liší svou schopností vychylovat segregační poměr ve svůj prospěch, a navíc se každý z nich může v přírodě vyskytovat ve více formách (alelách). Účinky jednotlivých genů se sčítají, a různé kombinace se proto liší „silou“ meiotického tahu a vlivem na plodnost či životaschopnost svých nositelů. 6) Kromě Tcd obsahuje t-haplotyp minimálně 16 recesivních letálních genů působících před narozením, které však nemají žádný vzájemný vztah a nemají ani nic společného s geny způsobujícími vychýlení transmisního poměru. Všeobecně se má za to, že vznik těchto genů (resp. jejich letálních účinků) byl podporován přírodním výběrem proto, že brání situaci, kdy neplodní a z biologického hlediska tedy nepotřební samci, homozygotní pro t-haplotyp, konkurují ostatním jedincům v soutěži o potravní zdroje. Zvýšená úmrtnost a snížená plodnost u homozygotů jsou důvodem, proč celý komplex rychle nevytlačí původní formu z populace, ale naopak je v ní udržován v překvapivě nízké frekvenci 5–15 %. A konečně, přestože je celý t-haplotyp díky paracentrickým inverzím „uzamčen“ do jednoho celku, ojedinělé crossing-overy, většinou mezi sousedními inverzemi (zhruba ve frekvenci 2–5 na 1000 pohlavních buněk), dávají vzniknout „částečným“ t-haplotypům, které si uchovávají pouze určitou část vlastností „kompletního“ komplexu.
Ultrasobecký element to nemá jednoduché
Život ultrasobeckého elementu není tak růžový, jak by se na první pohled mohlo zdát. Podobně jako u octomilky vyvinuly se i u myši domácí z některých genů na „normálním“ 17. chromozomu i na chromozomech dalších modifikátory (supresory), které se snaží nepříznivému působení t-haplotypů zabránit nebo ho alespoň zmírnit. Tak se t-haplotypy nacházejí uprostřed neustálé „chromozomální války“ (viz rámeček). Je zřejmé, že děje na úrovni jednotlivých genů mohou být stejně zajímavé jako vzájemné chování organizmů.Poznámky
-haplotypů
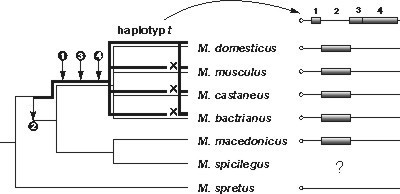 Problém původu t-haplotypů se zkoumá. Shromážděné údaje nejen naznačují odpovědi na otázky, kdy, kde, jak a proč tato struktura vznikla, ale vrhají nové světlo i na naše představy o jejich podstatě a evolučních mechanizmech, které je formují.
Problém původu t-haplotypů se zkoumá. Shromážděné údaje nejen naznačují odpovědi na otázky, kdy, kde, jak a proč tato struktura vznikla, ale vrhají nové světlo i na naše představy o jejich podstatě a evolučních mechanizmech, které je formují.
Klíčovou roli ve zkoumání původu t-haplotypů hrají dva úseky DNA tohoto komplexu, intron genu Tcp-1 (introny jsou nekódující úseky eukaryotických genů, vmezeřené mezi úseky kódující, tzv. exony), umístěný v oblasti druhé paracentrické inverze (in(17)2, viz článek), a pseudogen Hba-ps4 v místě čtvrté inverze (in(17)4). Studiem těchto sekvencí dospěli vědci k důležitým poznatkům
Ke stažení
 Článek ve formátu PDF [312,5 kB]
Článek ve formátu PDF [312,5 kB]
O autorovi
Miloš Macholán
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
