










Padesát let ediakarské fauny?
Ediakarská fauna je asi nejslavnějším paleontologickým fenoménem tohoto století. V Čechách nebyla (zatím) nalezena.
Sporný je i termín fauna. Správně by měl být v uvozovkách, protože fauna znamená živočišstvo, ale o podstatu ediakarských organizmů se dosud vedou spory. Jak uvidíme dále, může jít o mnohobuněčné organizmy, ale také o prvoky, řasy, lišejníky i příslušníky zcela vymřelé říše bioty. Nálezy v Ediakaře z konce 40. let pokládal jejich objevitel R. C. Sprigg původně za spodnokambrické, tedy současné s faunami již tehdy známými z celého světa. Později (koncem 50. let) se prokázalo, že pocházejí z pozdně proterozoických (starohorních) usazenin, tj. z vrstev asi 640 až 620 milionů let starých. Poté byla podobná pozdně proterozoická společenstva nalezena na všech kontitnentech kromě Antarktidy; rozsáhlý materiál byl popsán z různých míst v Austrálii, z Namibie, Newfounlandu, Ruska a střední Anglie. Z nejbohatších lokalit byly již získány celkem desítky tisíc jedinců.
Medúzy, řasy apod.: Ediakarské organizmy se od naprosté většiny mladších živočišných fosilií odlišují na první pohled tím, že nejde o mineralizované skelety (krunýře, lastury, kosti). R. C. Sprigg se domníval, že našel otisky planktonních a nektonních (tj. větších živočichů s vyvinutými orgány pohybu) organizmů s měkkými těly, ve velké většině medúzy. Byl však zaskočen jejich vynikajícím zachováním: uvážíme-li, že těla mnohých medúz obsahují až 99 % mořské vody, jeví se šance na zachování v jakémkoliv sedimentačním prostředí jako mizivá. Jak uvidíme dále, je Spriggova otázka dosud velmi živá a snad klíčová pro pokrok v poznávání ediakarské fauny.
Další „ediakarská“ fosilie, nalezená v opuštěném lomu v Leicestershiru v Anglii, byla pokládána za řasu. Po jejím srovnání s ediakarskými jedinci, které v té době intenzivně studoval australský paleontolog M. F. Glaessner, její objevitel neochotně připustil, že by mohlo jít o živočicha.
Víceméně stranou pozornosti zůstalo dílo H. D. Pfluga z počátku 70. let, jenž zpracovával nálezy z Namibie a došel k závěru, že všechny tamější ediakarské taxony patří k jednomu, zcela vymřelému kmeni, který nazval Petalonamae. Nesprávné pochopení toponomie některých jedinců a idealizované popisy taxonů učinily Pflugovo životní dílo spíše problematickým; nestalo se základem pro další bádání. Předznamenalo však přístup několika současných autorů – tendenci (z různých důvodů, jak uvidíme dále, logickou) pokud možno „hodit všechny ediakarské taxony do jednoho pytle“.
Glaessnerova škola: Glaessner vycházel z předpokladu, že ediakarská fauna je složena převážně ze zástupců dosud žijících živočišných kmenů, tříd a řádů. Zpočátku byli hlavními argumenty pro tento názor předpokládaní láčkovci, hlavně medúzy a osmičetní koráli („ediakarské“ rody Rangea a Charnia), ale brzy se staly důležitějšími bilaterálně souměrné taxony Dickinsonia (obrázek) a Spriggina, jejichž morfologie byla velmi blízká některým žijícím „mnohoštětinatým červům“, polychétům (recentní rod Spinther). Přímé srovnání takto časově odlehlých taxonů ovšem nutně muselo hraničit s bagatelizováním problému možné konvergence. Tak není divu, že M. Wade (1972) pokládal spintheridy za skupinu polychétů, která se vývojově oddělila už v prekambriu, což je názor dnes (vzhledem k přínosu molekulární biologie k vykreslení hrubých rysů fylogeneze) již neudržitelný. 
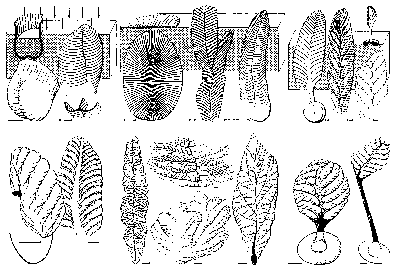
Vraťme se však k Seilacherově myšlence. Většina ediakarských organizmů měla ležet na dně, či být těsně zasypána sedimentem, nebo být zakotvena v sedimentu a spočívat na stvolu (obr. obrázek). Vlastní těla pak byla plochá, tvořená poměrně pevnou tělesnou stěnou a tekutinou vyplňující jednotlivé segmenty – masou protoplazmy vytvořenou fúzí měňavkovitých tělísek, aniž by došlo k fúzi jejich jader. Vzhledem k jedinečné konstrukci a další charakteristice vyplývající z hypotézy je nelze zařadit mezi živočichy, ani mezi houby, ani mezi rostliny, ani mezi bakterie či sinice; je tedy třeba vytvořit pro ně novou říší – Vendobionta.
Seilacher interpretuje některé další pozdně porterozoické fosilie jako primitivní láčkovce. Přítomnost zástupců dalších kmenů předpokládá též; ale měli skutečně měkká těla, nebyli schopni fosilizace a zanechali pouze stopy po činnosti – ichnofosilie.
Reakce na Seilacherovu myšlenku byla bouřlivá a vedla k explozi zájmu o ediakarské organizmy, která zatím – ani po více než deseti letech – nejeví známky umírnění, spíš naopak.
V devadesátých letech se autoři navazující víceméně na Glaessnerovu školu – převážně Australané: B. Runnegar, R.J.F. Jenkins aj. – snaží najít trhliny ve „vendobiontní hypotéze“ a naopak překlenout nedostatky hypotézy vlastní. Runnegar uvádí jako hlavní indicie proti Seilacherově hypotéze morfologickou podobnost některých ediakrských organizmů s mnohobuněčnými, malou pravděpodobnost současného objevení několika forem megaskopiského života (větších živočichů), přílišnou složitost některých struktur (dendritických kanálků u různých „vendobiontů“) na to, aby těla byla tvořena pouhou stěnou a „plazmodiálním fluidem“; v neposlední řadě pak poukazuje na nález „roztrženého“ jedince rodu Dickinsonia, jehož zachování je shodné jako u kusů „neroztržených“. Tělo bylo tedy tvořeno spíše buněčnou tkání než kapalinou, která by z poškozených segmentů vytekla.
Nově jsou popisovány známky (nutno říci, že velmi sporné) svalové aktivity a zbytky ústních otvorů u dickinsonií a spriggin.
Závěry pokračovatelů Glaessnerovy školy přijal např. S. Conway Morris, který učinil jeden z prvních pokusů o syntézu paleontologických a molekulárněbiologických dat při konstrukci „stromu života“. Téměř všechny ediakarské organizmy jsou do tohoto „stromu“ zabudovány jako mnohobuněční živočichové pestré taxonomické příslušnosti. Paleontologa možná udiví, že tento příspěvek uveřejnil bez kritické výhrady časopis z nejimpaktovanějších, zatímco nositel téměř Nobelovy ceny za geologii se závěry kategoricky nesouhlasí; další paleontologové s nepochybně vysokou citovaností pokládají pak domnělé mnohobuněčné za lišejníky, prvoky apod., jak uvidíme dále.
Trochu stranou Glaessnerovy školy stojí Rus M. A. Fedonkin, který zpracovával rozsáhlý a mnohdy jedinečný materiál z Ruska, Sibiře, Ukrajiny ad. Pokládá řadu ediakarských organizmů za členovce, zároveň však dokumentuje jejich morfologickou blízkost k radiálně symetrickým formám ediakarského života. Podle jeho názoru – bez ohledu na tvrzení opřená o poznatky molekulární biologie – docházelo k oddělování bilaterálně souměrných organizmů (např. arthropodů) od radiálně souměrných (např. láčkovců) právě během nejmladšího proterozoika. Molekulární biologové kladou oddělení těchto větví mnohem hlouběji do minulosti.
Uveďme ještě další zajímavé teorie o ediakarské fauně. Rus A. J. Žuravlev nejprve vyjmenovává důvody, proč by měly být ediakarské organizmy pokládány za jednolitý celek: společný půdorys, konstrukce a možnost nalézt plynulé morfologické přechody mezi „červy“, „členovci“, „láčkovci“ a dalšími (obr. obrázek). Dále pak zpochybňuje možnost zachování skutečně měkkých těl v usazeninách, které je možno pokládat za mělkovodní, okysličené, středně zrnité až hrubozrnné. Zmiňuje se o neúspěšných laboratorních pokusech s fosilizací medúz. Jediné jakžtakž „úspěšné“ byly Norrisovy pokusy otiskování do omítky. Jeho vlastní řešení? Nabízí srovnání s prvoky podkmene Xenophyophorida, kteří žijí přisedle v dnešních hlubokých mořích a jejich rozměry – ač jsou jednobuněční – odpovídají ediakarské fauně. Tito prvoci mají těla složená z plazmodia uzavřeného do systému větvících se trubic a z tuhých sraženin plazmodiem vyloučených. Sraženiny poskytují substrát pro symbiotickou mikroflóru, která je hlavním zdrojem potravy xenophyophorid. Žuravlev se brání přímému ztotožnění vendobiontů a xenophyophorid, ale přimlouvá se za zařazení ediakarských organizmů mezi prvoky. G. J. Retallack vystoupil s podobně koncipovanou teorií, že ediakarská fauna je tvořena lišejníky. 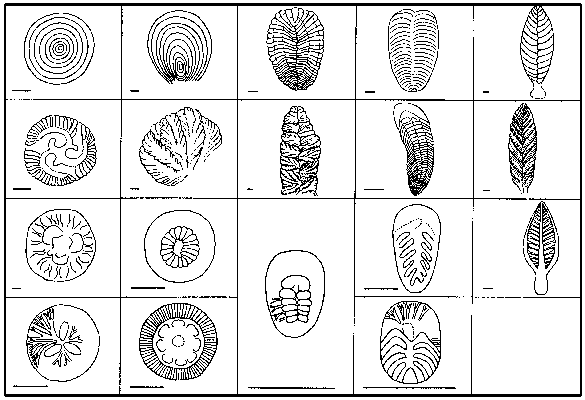
Ještě jeden současný myšlenkový směr je nutno zaznamenat. Je to snaha najít přímé potomky ediakarské „fauny“ ve spodním paleozoiku, jinými slovy dokázat, že ediakarská fauna s koncem starohor nevymřela úplně a naráz. Tak jsou s ediakarskými organizmy srovnávány některé prvky spodnokambrické fauny burgeských břidlic, severoevropská kambrická fosilie Xenusion aj. Zajímavé jsou nálezy, které popsal T. P. Crimes se spolupracovníky z hlubokomořských svrchnokambrických usazenin Irska. Jde o otisky diskovitých těles na spodních stranách vrstev, usazených gravitačními (turbiditními) proudy. Morfologie tělísek (obr. obrázek) je shodná s některými ediakarskými „láčkovci“ a jejich vysvětlení jako pseudofosilií není pravděpodobné. Pokud by přímé srovnání těchto organizmů s ediakarskou faunou bylo na místě, objasnily by se tím hned dva velké problémy: jednak že ediakarské organizmy přežily do paleozoika a dále že jejich těla byla pevná, protože jinak by se na bázi turbiditního proudu nemohla otisknout. 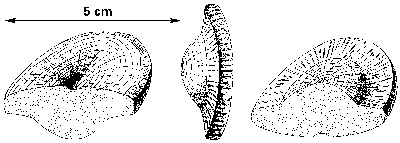
– v sedimentologických a tafonomických studiích (mohly by se spolehlivěji určit meze měkkosti, resp. pevnosti těl, některé vlastnosti stavebního materiálu aj.),
– v pokračujícím srovnávání paleontologického záznamu s důkazy přinášenými molekulární biologií (i když je jasné, že ribonukleovou kyselinu ediakarských organizmů nikdo nikdy neuvidí, je zde šance vyloučit alespoň některé konstrukce „stromu života“ jako chybné), – v relativně nové snaze najít přímé pokračovatele ediakarských taxonů i v paleozoiku.
Nálezy nových, lépe zachovalých fosilií na tradičních lokalitách nelze samozřejmě vyloučit, ale soudě podle množství kusů již nalezených bych řekl, že na to nelze ani příliš spoléhat. Je možné, že některé teorie se zhroutí poměrně rychle, ale lze si představit, že všechny současné směry zůstanou při životě i po dalších padesát let studia ediakarské „fauny“.
Tím, že při hledání ediakarských organizmů byla překročena hranice proterozoika a paleozoika, vyvstává možnost nálezů ediakarské fauny i u nás. Žádný paleontolog pracující v barrandienu by asi nesouhlasil s tím, že tam už byli nalezeni nějací vendobionti, jen nebyli správně interpretováni. A přesto nám to bylo již nejméně jednou (byť mimopublikačně) podsouváno. A může se to opakovat. To je také hlavní důvod, proč jsem se během stáže na jednom z pracovišť, kde je ediakarská fauna zkoumána, pustil do hromady literatury o problému. Šance na objevení tak významné (a dobře prodejné) tematiky na domácí scéně je sice vítána, avšak zatažení do často vágních diskusí opřených o povrchní morfologickou podobnost by mohli někteří příslušníci naší paleontologické veřejnosti vnímat nelibě, jako laciný humbuk.Literatura
Glaessner M. F.: Precambrian Coelenterata from Australia, Africa and England, Nature 183, 1472 – 1473, 1959Glaessner M. F.: Pre-Cambrian animals, Scientific American 204, 72 –78, 1961
Seilacher A.: Precambrian metazoan extinctions, Abstr. Prog. Geol. Soc. Amer. 15, 683, 1983
Seilacher A.: Vendobionta and Psammocorallia: lost constructions of Precambrian evolution, J. Geol. Soc. 149, 607 – 613, London 1992
Retallack G. J.: Were the Ediacaran fossils lichenlike organism? Geol. Soc. Amer. Abstract with Programs 24, A226 – A227, 1992
Conway Morris S.: The fossil record and the early evolution of Metazoa, Nature 361, 219 – 225, 1993
Crimes T. P., Insole A., Williams B. P. J.: A rigid-bodied Ediacaran Biota from Upper Cambrian strata in Co. Wexford, Eire. Geol. J. 30, 89 – 109, Liverpool 1995
PALEONTOLOGIE MODERNĚ POJATÁ?
Spojené státy jsou podobně jako Česká republika klasickou zemí paleontologie. Útlum aktivit spojených s vyhledáváním nových ložisek nerostných surovin však způsobil, že řada paleontologů přešla na pracoviště zabývající se biologickou problematikou. Profesorka Marvalee Wake, vedoucí oddělení integrativní biologie na Kalifornské univerzitě v Berkeley, k tomu říká:
Po rozsáhlé reorganizaci biologických věd na Kalifornské univerzitě v Berkeley koncem osmdesátých let je paleontologie začleněna do oddělení tzv. integrativní biologie. Pod tímto názvem jsou zahrnuty následující disciplíny: biologie organizmů, ekologie, etologie, evoluce, systematika, morfologie, fyziologie, biomechanika a endokrinologie. Pracoviště sdružuje pracovníky bývalých oddělení zoologie, botaniky, paleontologie, anatomie a fyziologie, a rovněž oddělení genetiky (toto poslední oddělení se při reorganizaci rozdělilo, část přešla k oddělení integrativní biologie, část k oddělení molekulární a buněčné biologie). Oddělení paleontologie bylo před reorganizací posledním paleontologickým pracovištěm na významnějších univerzitách ve Spojených státech, které si udrželo organizační nezávislost, a proto pracovníci tohoto oddělení zpočátku projevovali k reorganizaci nedůvěru. Všichni se však posléze shodli na tom, že paleontologie na univerzitě v Berkeley se již v minulosti orientovala výrazně biologickým směrem (evoluce, systematika, paleoekologie, morfologie, biomechanika, se zaměřením na fosilní prvoky, rostliny i živočichy) a že těsná spolupráce mezi paleontology a neontology, kteří se zabývají stejnými základními problémy, může řešení těchto problémů zefektivnit. Podle našeho názoru by se nemělo řešení těchto otázek svazovat jen s určitými taxony (vymřelými či recentními) nebo jen s určitými časovými úseky (minulost, recent). K oddělení integrativní biologie bylo přičleněno i paleontologické muzeum, protože paleontologie potřebuje rozsáhlou databázi v podobě fosilií. Muzeum si však zachovává administrativní nezávislost a tím se do značné míry udržuje kontinuita samostatné paleontologie na naší univerzitě. Podobně je přičleněno i zoologické muzeum obratlovců a dva rozsáhlé herbáře. Tyto sbírkové fondy jsou společně umístěny v nové budově v bezprostřední blízkosti našeho oddělení, což usnadňuje jejich vědecké a pedagogické využití.
O autorovi
Radek Mikuláš

Doporučujeme

