









Živoucí kameny mezi lišejníky
| 9. 10. 2014Mnohá jsoucna vznikají v přírodě nenáhodně a opakovaně. Přestože vykazují variabilitu, není většinou těžké odhalit, že jde o variaci na totéž. Většina známých lišejníků má pletiva ve stélce upořádaná v souvislých, s povrchem rovnoběžných vrstvách. Některé vzájemně si nepříbuzné druhy však vytvářejí vysoké „světlovody“ z houbových hyf, které jsou obklopeny sloupci z řasových buněk. Troufáme si tvrdit, že tuto pozoruhodnou reorganizaci stélek některých lišejníků vyprovokovaly podobné ekologické potřeby, které vedly ke vzniku „živých kamenů“ mezi sukulenty.
Živé kameny
Kaktusáři a nejen oni je znají – sukulenty převážně jihoafrických pouští, zčásti nebo celé pod povrchem půdy (nejznámější je patrně rod Lithops). Vykukuje jim často jen šedá ploška, která připomíná kámen (odtud jméno). Při bližším prozkoumání zjistíme, že jde o průsvitný povrch stonku či listu, o jakési okénko do bizarní podzemní rostliny. Pod okénkem se nachází bezbarvé pletivo propouštějící světlo k zeleným fotosyntetizujícím buňkám po obvodu a ve spodní části těla rostliny (obr. 1). Fotosyntéza tak může probíhat za příznivějších podmínek než za spalujícího žáru panujícího na povrchu půdy. Rostlina pak také zřejmě lépe hospodaří s vodou.
 Přestože několik prací studujících (eko)fyziologický význam listových či stonkových okének sukulentů přineslo poněkud rozporuplná zjištění, přínos této morfologie je nesporný. Vyskytuje se nezávisle u několika nepříbuzných čeledí sukulentních rostlin (nejlépe je vyvinuta u kosmatcovitých, liliovitých a hvězdnicovitých). Je tedy jisté, že v evoluci vznikla nezávisle a vícekrát. „Okénkaté sukulenty“ se navíc vyskytují v oblastech s vysokými teplotami a extrémní sluneční ozářeností ve vegetačních měsících.
Přestože několik prací studujících (eko)fyziologický význam listových či stonkových okének sukulentů přineslo poněkud rozporuplná zjištění, přínos této morfologie je nesporný. Vyskytuje se nezávisle u několika nepříbuzných čeledí sukulentních rostlin (nejlépe je vyvinuta u kosmatcovitých, liliovitých a hvězdnicovitých). Je tedy jisté, že v evoluci vznikla nezávisle a vícekrát. „Okénkaté sukulenty“ se navíc vyskytují v oblastech s vysokými teplotami a extrémní sluneční ozářeností ve vegetačních měsících.
I ty, Brute?
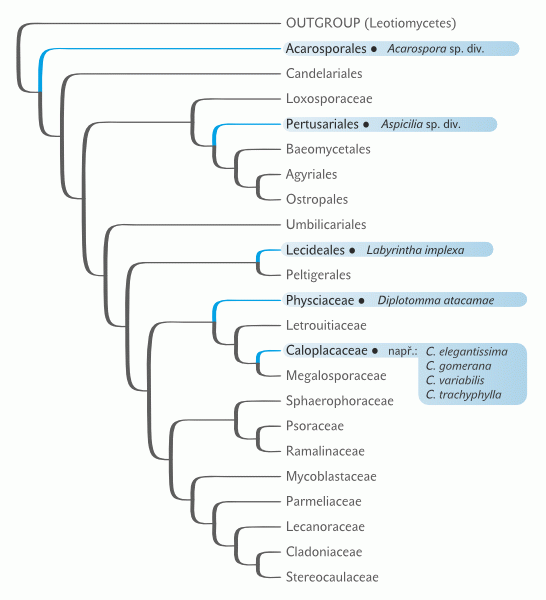
Morfologie či funkce, které jsou v daném prostředí dlouhodoběji výhodné, vznikají opakovaně, a to často dosti odlišnými evolučními cestičkami. To není nic nového. Aktivně létat se naučil hmyz, ptáci (původně plazi) a savci. Protože využití vzdušné ekologické niky létáním se prostě vyplácí. C4 a CAM fotosyntéza (Vesmír 80, 43, 2001/1; 91, 35, 2012/1) vznikly v evoluci cévnatých rostlin mnohokrát nezávisle, protože v jisté kombinaci faktorů jsou výhodné. Možná proto nepřekvapí, že okénka a světlovodná pletiva najdeme i jinde než u rostlin. Tato anatomie vznikla vícekrát nezávisle i mezi lišejníky (obr. 2 a 3). Mnohočetný vznik znaku či funkce v evoluci skoro vždy ukazuje na společný ekologický tlak a kompetiční výhodu, kterou tato inovace v daném prostředí a čase přináší.
Kde rostou „lišejníkové živoucí kameny“?
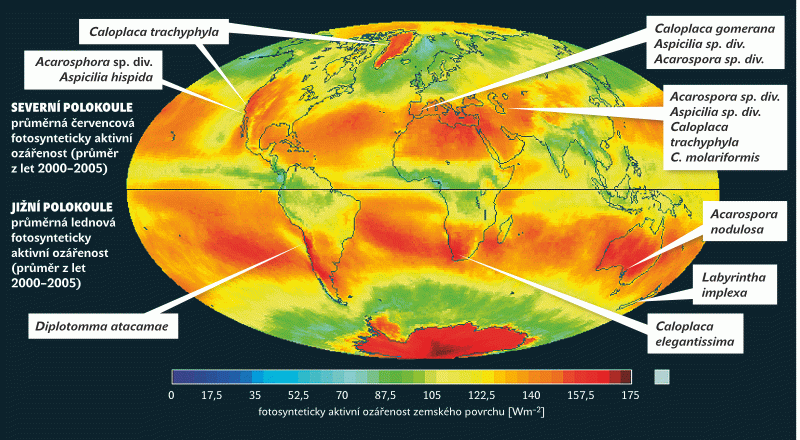
Ekologická nika je mnohorozměrný časoprostor vymezený všemi faktory prostředí, které na daný druh působí. Není proto mnohdy snadné říci, kde jsou hranice takové niky, proč daná adaptace (zde a nyní) přináší výhodu a v čem přesně spočívá. Jistě není snadné odhalit, proč se z pohyblivých živočichů nenaučilo více skupin létat. Vždyť létání musí být přece tak výhodné (kolik jen hudebních skupin pojmenovalo své písně „If I could fly“!). O to překvapivější je, že se podařilo najít jeden faktor prostředí, v jehož maximu se – s jednou výjimkou – nachází těžiště výskytu všech známých „okénkatých lišejníků“. Tímto faktorem je osluněnost (insolace) v nejletnějším měsíci (červenec na severní a leden na jižní polokouli).
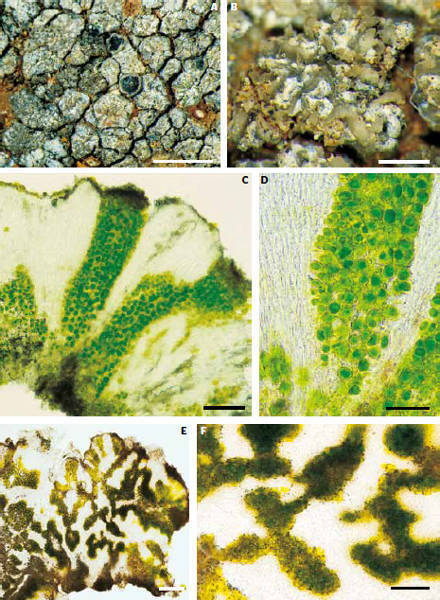 Krom jednoho druhu všechny dosud známé lišejníky se světlovodnými pletivy kopírují velmi přesně svým výskytem oblasti s nejvyšší osluněností (cca 160–180 W/m2; obr 4). Tou jednou výjimkou je druh Labyrintha implexa, který je endemitem Nového Zélandu. Maxima insolace jsou zde sice nadprůměrná (120–140 W/m2), ale nikoli extrémní. Výjimka může potvrzovat pravidlo. Navíc musíme zohlednit teorii ostrovní speciace a pohybu pevnin. V geologické minulosti, kdy ke vzniku této morfologie došlo (neví se přesně, kdy to bylo), mohl ostrov ležet jinde (nebo být součástí pevniny) a podmínky mohly být odlišné. Zajímavé je, že právě u druhu Labyrintha implexa je světlovodná morfologie vyvinuta vůbec nejlépe ze známých druhů. Toho si povšiml W. Malcolm, optik a lichenolog. Měřil prostup světla houbovými světlovodnými pletivy vlhkých, tedy aktivních lišejníků. Zjistil, že prostup světla je velmi vysoký a významné procento světla se dostává až k řasovým buňkám při bázi lišejníkové stélky na kontaktu se substrátem.
Krom jednoho druhu všechny dosud známé lišejníky se světlovodnými pletivy kopírují velmi přesně svým výskytem oblasti s nejvyšší osluněností (cca 160–180 W/m2; obr 4). Tou jednou výjimkou je druh Labyrintha implexa, který je endemitem Nového Zélandu. Maxima insolace jsou zde sice nadprůměrná (120–140 W/m2), ale nikoli extrémní. Výjimka může potvrzovat pravidlo. Navíc musíme zohlednit teorii ostrovní speciace a pohybu pevnin. V geologické minulosti, kdy ke vzniku této morfologie došlo (neví se přesně, kdy to bylo), mohl ostrov ležet jinde (nebo být součástí pevniny) a podmínky mohly být odlišné. Zajímavé je, že právě u druhu Labyrintha implexa je světlovodná morfologie vyvinuta vůbec nejlépe ze známých druhů. Toho si povšiml W. Malcolm, optik a lichenolog. Měřil prostup světla houbovými světlovodnými pletivy vlhkých, tedy aktivních lišejníků. Zjistil, že prostup světla je velmi vysoký a významné procento světla se dostává až k řasovým buňkám při bázi lišejníkové stélky na kontaktu se substrátem.
Ekofyziologie lišejníků se světlovodnou morfologií
Světlovodné uspořádání houbových vláken tedy vzniklo vícekrát nezávisle v evoluci lišejníkové symbiózy a vyskytuje se v oblastech s extrémním osluněním. Je tedy logické se ptát, jaké funkční vlastnosti lišejníků stojí za touto ekologickou úspěšností a „opakovaným vstupem do stejné evoluční řeky“.
Na začátku našeho výzkumu stály dvě hypotézy:
 1. Lišejníkové živoucí kameny díky vyšší tloušťce (obr. 2) vysychají pomaleji než ostatní lišejníky. Pro lišejníky pouštních a stepních oblastí je doba aktivity klíčová pro fotosyntézu – příjem CO2, který je nezbytný pro růst. Když slunce vyjde nad obzor, lišejníky aridních oblastí jsou většinou již nějakou dobu aktivní (vlhké) díky ranní rose. Ostré slunce je však většinou vysuší během hodiny, málokdy několika hodin. Zbytek dne přečkávají v neaktivním suchém stavu a čekají na rosu, která je příští noci opět aktivuje. I nevelké prodloužení aktivity (během světlé části dne) může být důležité. Světlo je po rozbřesku stále intenzivnější, většinu však lišejníky nedokážou využít – jsou už suché. Pomalejší vysychání lišejníkových živoucích kamenů vůči ostatním lišejníkům jsme však v laboratorních podmínkách neprokázali. To ovšem neznamená, že se v přírodních podmínkách neprojevuje.
1. Lišejníkové živoucí kameny díky vyšší tloušťce (obr. 2) vysychají pomaleji než ostatní lišejníky. Pro lišejníky pouštních a stepních oblastí je doba aktivity klíčová pro fotosyntézu – příjem CO2, který je nezbytný pro růst. Když slunce vyjde nad obzor, lišejníky aridních oblastí jsou většinou již nějakou dobu aktivní (vlhké) díky ranní rose. Ostré slunce je však většinou vysuší během hodiny, málokdy několika hodin. Zbytek dne přečkávají v neaktivním suchém stavu a čekají na rosu, která je příští noci opět aktivuje. I nevelké prodloužení aktivity (během světlé části dne) může být důležité. Světlo je po rozbřesku stále intenzivnější, většinu však lišejníky nedokážou využít – jsou už suché. Pomalejší vysychání lišejníkových živoucích kamenů vůči ostatním lišejníkům jsme však v laboratorních podmínkách neprokázali. To ovšem neznamená, že se v přírodních podmínkách neprojevuje.
2. Lišejníkové živoucí kameny dosahují vyšší intenzity fotosyntézy než ostatní lišejníky. Lišejníky se světlovodnou morfologií bývají mnohem tlustší než většina korovitých lišejníků (obr. 2). Připomínají tak slunné listy (např. u buku). Slunné listy mnohých druhů jsou zhruba 2× silnější než listy stinné. Zároveň slunné listy mají vrstvu dlouhých, tzv. palisádových buněk, jejichž světlovodná funkce byla již dříve prokázána. Ale především: slunné listy mají vyšší maximální fotosyntézu (rychlost příjmu oxidu uhličitého) než listy stinné. A fotosyntéza (částečně) určuje rychlost růstu a rychlost růstu pak konkurenceschopnost. Tato hypotéza se u našich vzorků z Kazachstánu potvrdila (obr. 5).
Lišejníkové živoucí kameny využívají silné světlo účinněji než ostatní korovité lišejníky
 S výjimkou jednoho druhu měly „živoucí kameny mezi lišejníky“ při silném světle vyšší příjem oxidu uhličitého než druhy s řasovými buňkami rozmístěnými v horizontální souvislé vrstvě. To vede k rychlejšímu příjmu CO2, a tím i k nárůstu biomasy za vysoké osluněnosti. Zároveň však tyto druhy vykazovaly vyšší dýchání ve tmě. Dýchání, ve výsledku opačný proces než je fotosyntéza, vede ke ztrátě uhlíku a biomasy. Dýchání převládá za nedostatku světla, ale také v jinak nepříznivých podmínkách (např. nedostatečná hydratace). Je tedy zřejmé, že bude existovat průsečík podmínek, kdy se vyplatí být „živoucím kamenem“, a kdy naopak lišejníkem s běžnou anatomickou stavbou. Tento průsečík se týká hlavně úhrnného oslunění v čase, kdy je lišejník aktivní. Pokud druh roste v zástinu vyšších pater vegetace nebo tam, kde slunce kvůli časté oblačnosti moc nesvítí, je výhodné mít klasickou vrstevnatou anatomii, a tedy nižší dýchání. Naopak v místech, kde slunce svítívá už od rozbřesku a kde nejsou lišejníky zastíněny, budou výhodné „světlovody“ lišejníkových živoucích kamenů, které umožní intenzivnější asimilaci oxidu uhličitého.
S výjimkou jednoho druhu měly „živoucí kameny mezi lišejníky“ při silném světle vyšší příjem oxidu uhličitého než druhy s řasovými buňkami rozmístěnými v horizontální souvislé vrstvě. To vede k rychlejšímu příjmu CO2, a tím i k nárůstu biomasy za vysoké osluněnosti. Zároveň však tyto druhy vykazovaly vyšší dýchání ve tmě. Dýchání, ve výsledku opačný proces než je fotosyntéza, vede ke ztrátě uhlíku a biomasy. Dýchání převládá za nedostatku světla, ale také v jinak nepříznivých podmínkách (např. nedostatečná hydratace). Je tedy zřejmé, že bude existovat průsečík podmínek, kdy se vyplatí být „živoucím kamenem“, a kdy naopak lišejníkem s běžnou anatomickou stavbou. Tento průsečík se týká hlavně úhrnného oslunění v čase, kdy je lišejník aktivní. Pokud druh roste v zástinu vyšších pater vegetace nebo tam, kde slunce kvůli časté oblačnosti moc nesvítí, je výhodné mít klasickou vrstevnatou anatomii, a tedy nižší dýchání. Naopak v místech, kde slunce svítívá už od rozbřesku a kde nejsou lišejníky zastíněny, budou výhodné „světlovody“ lišejníkových živoucích kamenů, které umožní intenzivnější asimilaci oxidu uhličitého.
A je zde ekofyziologie opravdu určující?
Fyziologická šetření přinášejí cenné informace o fungování konkrétních tvorů. Bohužel, v přírodě je situace často tak složitá (mnoho kombinujících a vzájemně se ovlivňujících faktorů), že je velmi obtížné změřit všechny vlivy prostředí, modelovat je a extrapolovat v ekologickou úspěšnost druhu.
 Lišejníky často rostou na místech, kde byliny, keře, stromy a často ani mechy nepřežijí. To ovšem neznamená, že si vzájemně nekonkurují a nebojují mezi sebou. Když se podíváme na mnohé skalní podklady, které jsou téměř souvisle pokryty stélkami lišejníků, mělo by nám to být jasné. I lišejníky tu musí mezi sebou soupeřit. A právě vzájemné vztahy spolu žijících druhů jsou často pro jejich přežívání důležitější než ty vybrané fyziologické vlastnosti, kterých si povšimneme a které dokážeme správně měřit. Tak například následující, zdánlivě triviální zákonitost může existenci lišejníkových živoucích kamenů elegantně vysvětlovat i bez fyziologického zkoumání. Houbové světlovody lišejníkům umožňují, aby narostly do vyšší tloušťky než lišejníky s běžnou morfologií. Pozorování v přírodě jasně ukazují, že tlustší stélky běžně přerůstají ty tenčí. Naopak jsme nepozorovali, že by tenčí stélky konkurenčně ohrožovaly ty tlustší. Náš současný výzkum je zaměřen právě na pochopení omezeného geografického rozšíření konkurenčně silných, tlustých lišejníkových krust.
Lišejníky často rostou na místech, kde byliny, keře, stromy a často ani mechy nepřežijí. To ovšem neznamená, že si vzájemně nekonkurují a nebojují mezi sebou. Když se podíváme na mnohé skalní podklady, které jsou téměř souvisle pokryty stélkami lišejníků, mělo by nám to být jasné. I lišejníky tu musí mezi sebou soupeřit. A právě vzájemné vztahy spolu žijících druhů jsou často pro jejich přežívání důležitější než ty vybrané fyziologické vlastnosti, kterých si povšimneme a které dokážeme správně měřit. Tak například následující, zdánlivě triviální zákonitost může existenci lišejníkových živoucích kamenů elegantně vysvětlovat i bez fyziologického zkoumání. Houbové světlovody lišejníkům umožňují, aby narostly do vyšší tloušťky než lišejníky s běžnou morfologií. Pozorování v přírodě jasně ukazují, že tlustší stélky běžně přerůstají ty tenčí. Naopak jsme nepozorovali, že by tenčí stélky konkurenčně ohrožovaly ty tlustší. Náš současný výzkum je zaměřen právě na pochopení omezeného geografického rozšíření konkurenčně silných, tlustých lišejníkových krust.
Literatura
Malcolm W. M.: Light transmission inside the thallus of Labyrintha implexa (Porpidiaceae, lichenized Ascomycetes). Bibliotheca Lichenologica 58, 275–280, 1995.
Vondrák J., Kubásek J.: Algal stacks and fungal stacks as a adaptation to high light in lichens. Lichenologist 45, 115–124, 2013.
Ke stažení
 článek ve formátu pdf [1,13 MB]
článek ve formátu pdf [1,13 MB]
O autorech
Jiří Kubásek
Jan Vondrák
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
