









Mnohobuněčné bakterie a jejich projevy
| 3. 5. 2012 Všichni tvorové v biosféře jsou do různé míry vnořeni do složité a proměnlivé sítě ekologických (symbiotických) vztahů; na jedné straně síť neustále moduluje jejich životaběh, na straně druhé sama jejich existence pozměňuje síť. Ontogeneze čili utváření mnohobuněčného, druhově specifického těla z jediné buňky nebo skupiny blízce příbuzných buněk (např. u hlenek) bývá téměř vždy silně ovlivněna přítomností jiných bytostí. Proto je např. ontogeneze amniotických obratlovců zpočátku zcela izolována od zbytku biosféry: embryogeneze a raný vývoj plodu probíhají ve sterilním prostředí. Novorozený jedinec se napojí na biosférickou síť až v pokročilém stadiu vývoje. Výzkum bezmikrobních neboli gnotobiotických zvířat (často i rostlin) odhaluje nesmírné obtíže, které musí jedinec překonat, žije-li i po narození ve sterilním nebo jen polosterilním prostředí. Uměle navozený stav gnotobiózy nám však pomůže zahlédnout některé vztahy (např. mezi makroorganismem a symbionty jeho útrob nebo mezi symbionty samotnými), které by se na pozadí stovek jiných interakcí ztrácely.
Všichni tvorové v biosféře jsou do různé míry vnořeni do složité a proměnlivé sítě ekologických (symbiotických) vztahů; na jedné straně síť neustále moduluje jejich životaběh, na straně druhé sama jejich existence pozměňuje síť. Ontogeneze čili utváření mnohobuněčného, druhově specifického těla z jediné buňky nebo skupiny blízce příbuzných buněk (např. u hlenek) bývá téměř vždy silně ovlivněna přítomností jiných bytostí. Proto je např. ontogeneze amniotických obratlovců zpočátku zcela izolována od zbytku biosféry: embryogeneze a raný vývoj plodu probíhají ve sterilním prostředí. Novorozený jedinec se napojí na biosférickou síť až v pokročilém stadiu vývoje. Výzkum bezmikrobních neboli gnotobiotických zvířat (často i rostlin) odhaluje nesmírné obtíže, které musí jedinec překonat, žije-li i po narození ve sterilním nebo jen polosterilním prostředí. Uměle navozený stav gnotobiózy nám však pomůže zahlédnout některé vztahy (např. mezi makroorganismem a symbionty jeho útrob nebo mezi symbionty samotnými), které by se na pozadí stovek jiných interakcí ztrácely.
 Na opačném pólu ontogenetických vztahů jsou složitá společenstva složená ze stovek druhů. Teprve jejich vzájemná souhra umožní vznik společenstva, jakým je mikrobiom zubního povlaku či rybničního sedimentu, která zřídka mají jasně rozpoznatelný makroskopický tvar; jejich propracované a často filigránské uspořádání se projeví teprve na mikroskopické a biochemické úrovni. Mikroorganismy zúčastňující se této hry jen málokdy dostanou příležitost vytvořit mnohobuněčné tělo zcela či aspoň zčásti izolované od zbytku sítě. Obvykle k tomu dochází na čerstvě dostupných bohatých substrátech (puklá hruška, povrch smetany anebo umělé podmínky v laboratoři). Tam pak pozorujeme nádheru bakteriálních či plísňových kolonií. I tak je ontogeneze velmi plastická – snadno ovlivnitelná drobnými obměnami kultivačního prostředí (teploty, složení živin, hutnosti substrátu ap.).
Na opačném pólu ontogenetických vztahů jsou složitá společenstva složená ze stovek druhů. Teprve jejich vzájemná souhra umožní vznik společenstva, jakým je mikrobiom zubního povlaku či rybničního sedimentu, která zřídka mají jasně rozpoznatelný makroskopický tvar; jejich propracované a často filigránské uspořádání se projeví teprve na mikroskopické a biochemické úrovni. Mikroorganismy zúčastňující se této hry jen málokdy dostanou příležitost vytvořit mnohobuněčné tělo zcela či aspoň zčásti izolované od zbytku sítě. Obvykle k tomu dochází na čerstvě dostupných bohatých substrátech (puklá hruška, povrch smetany anebo umělé podmínky v laboratoři). Tam pak pozorujeme nádheru bakteriálních či plísňových kolonií. I tak je ontogeneze velmi plastická – snadno ovlivnitelná drobnými obměnami kultivačního prostředí (teploty, složení živin, hutnosti substrátu ap.).
 Naše skupina studuje ontogenezi kolonií Serratia sp. za umělých bezmikrobních podmínek. Ukázali jsme, jak na bohatých půdách dokáže tento „mikroorganismus“ rozehrát svůj tvarový potenciál. Studovali jsme „gnotobiologické“ vztahy mezi koloniemi jednoho druhu, anebo blízce příbuznými klony odvozenými ze stejného mateřského izolátu (vzájemné ovlivňování v čase a prostoru). Pracovali jsme s pěti stabilními a snadno rozpoznatelnými morfotypy bakteriálních druhů Serratia rubidaea, Serratia marcescens a Escherichia coli. Jde vždy o gramnegativní fakultativně anaerobní organismy, které spadají do stejné taxonomické skupiny Enterobacteriae. Náš kmen S. rubidaea produkuje dva stabilní typy kolonií (morfotypy) – původní morfotyp R s hladkými červenými koloniemi o průměru 20 mm a dceřiný bílý morfotyp W (obr. 1A vlevo). S. marcescens vytváří také dva morfotypy, které jsme podle profilu stavby kolonií nazvali „fontánky“ – původní červenobílý (F) a z něho odvozený bílý Fw (obr. 1B vlevo). Fontánkové kolonie dorůstají průměru 15 mm a skládají se z vyvýšeného středu, mezikruží a lemu. Dceřiné bílé kolonie se v obou případech od svých mateřských morfotypů R a F kromě zbarvení neliší. Kolonie všech uvedených morfotypů vykazují konečný růst a jejich vzhled není závislý na způsobu sázení, pokud použijeme jeden z těchto tří typů: klonální růst z jedné buňky, vykapání husté suspenze na malou plochu (o průměru menším než 1 mm), nebo „bodové“ nanesení materiálu odebraného z kolonie. Spolu s výraznými barevnými projevy tím získáme dobře definovaný materiál ke studiu morfogeneze i vztahů koloniálních jedinců v jednoduchém „ekosystému“ misky.
Naše skupina studuje ontogenezi kolonií Serratia sp. za umělých bezmikrobních podmínek. Ukázali jsme, jak na bohatých půdách dokáže tento „mikroorganismus“ rozehrát svůj tvarový potenciál. Studovali jsme „gnotobiologické“ vztahy mezi koloniemi jednoho druhu, anebo blízce příbuznými klony odvozenými ze stejného mateřského izolátu (vzájemné ovlivňování v čase a prostoru). Pracovali jsme s pěti stabilními a snadno rozpoznatelnými morfotypy bakteriálních druhů Serratia rubidaea, Serratia marcescens a Escherichia coli. Jde vždy o gramnegativní fakultativně anaerobní organismy, které spadají do stejné taxonomické skupiny Enterobacteriae. Náš kmen S. rubidaea produkuje dva stabilní typy kolonií (morfotypy) – původní morfotyp R s hladkými červenými koloniemi o průměru 20 mm a dceřiný bílý morfotyp W (obr. 1A vlevo). S. marcescens vytváří také dva morfotypy, které jsme podle profilu stavby kolonií nazvali „fontánky“ – původní červenobílý (F) a z něho odvozený bílý Fw (obr. 1B vlevo). Fontánkové kolonie dorůstají průměru 15 mm a skládají se z vyvýšeného středu, mezikruží a lemu. Dceřiné bílé kolonie se v obou případech od svých mateřských morfotypů R a F kromě zbarvení neliší. Kolonie všech uvedených morfotypů vykazují konečný růst a jejich vzhled není závislý na způsobu sázení, pokud použijeme jeden z těchto tří typů: klonální růst z jedné buňky, vykapání husté suspenze na malou plochu (o průměru menším než 1 mm), nebo „bodové“ nanesení materiálu odebraného z kolonie. Spolu s výraznými barevnými projevy tím získáme dobře definovaný materiál ke studiu morfogeneze i vztahů koloniálních jedinců v jednoduchém „ekosystému“ misky.
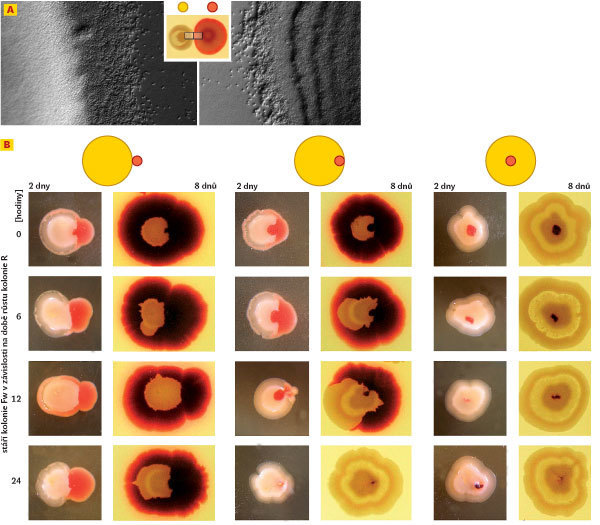 Vývoj jednotlivých morfotypů a vzájemné rozdíly mezi hladkými koloniemi S. rubidaea (R, W) a fontánkovými koloniemi S. marcescens (F, Fw) lze pozorovat nejenom makroskopicky, ale též na mikroskopických detailech vývinu jejich okrajů. V době růstu (zhruba do 8. dne vývoje) vysílají kolonie volné „skautující“ buňky do okolního prostoru. Výraznou odlišností u hladkých morfotypů je tvorba okrajových kaskádových teras, které u fontánkových morfotypů chybějí. Tyto rozdíly zůstávají zachovány i po ukončení růstu, poté co plně vyvinuté kolonie ukončí vysílání skautů (obr. 1A, B vpravo). Význam skautujících buněk zatím není úplně jasný. Pozoruhodnou záležitostí ve vývoji morfotypů F je vznik struktury X – přídatného bílého lemu po obvodu celé kolonie (obr. 2A), který je typickou reakcí fontánkových kolonií na přítomnost kolonie jiné než fontánkové. Kolonie F takto reaguje na souseda v jakémkoliv stadiu svého vývoje, jako kdyby se vůči němu vymezovala a izolovala. Po celém obvodu je lemována „aurou“ skautujících buněk (obr. 2B), na rozdíl od plně vyvinuté kolonie F. Tato změna je vývojová, nikoliv genetická. Po odebrání buněčného materiálu ze struktury X a opětovném vysetí dostáváme standardní fontánkové potomstvo.
Vývoj jednotlivých morfotypů a vzájemné rozdíly mezi hladkými koloniemi S. rubidaea (R, W) a fontánkovými koloniemi S. marcescens (F, Fw) lze pozorovat nejenom makroskopicky, ale též na mikroskopických detailech vývinu jejich okrajů. V době růstu (zhruba do 8. dne vývoje) vysílají kolonie volné „skautující“ buňky do okolního prostoru. Výraznou odlišností u hladkých morfotypů je tvorba okrajových kaskádových teras, které u fontánkových morfotypů chybějí. Tyto rozdíly zůstávají zachovány i po ukončení růstu, poté co plně vyvinuté kolonie ukončí vysílání skautů (obr. 1A, B vpravo). Význam skautujících buněk zatím není úplně jasný. Pozoruhodnou záležitostí ve vývoji morfotypů F je vznik struktury X – přídatného bílého lemu po obvodu celé kolonie (obr. 2A), který je typickou reakcí fontánkových kolonií na přítomnost kolonie jiné než fontánkové. Kolonie F takto reaguje na souseda v jakémkoliv stadiu svého vývoje, jako kdyby se vůči němu vymezovala a izolovala. Po celém obvodu je lemována „aurou“ skautujících buněk (obr. 2B), na rozdíl od plně vyvinuté kolonie F. Tato změna je vývojová, nikoliv genetická. Po odebrání buněčného materiálu ze struktury X a opětovném vysetí dostáváme standardní fontánkové potomstvo.
Kolonie ve vzájemném styku
Výsledná morfologie bakteriálních kolonií je velmi citlivá ke změnám prostředí. Kolonie reaguje na změny abiotické (teplota, pH, složení média atd.), ale neméně významnou roli při morfogenezi hraje i přítomnost jiného bakteriálního těla. Může jít o souseda stejného (interakce homospecifické) nebo i jiného (interakce heterospecifické) druhu.
 Při zkoumání homospecifického působení dvou morfotypů jsme sázeli kolonie v různých vzdálenostech od sebe. Vzdálenost mezi sousedy v podstatě simuluje stupeň vývoje, při němž se kolonie setkají. Pokud se setkají mladší vývojová stadia, mohou se vzájemně dotknout, eventuálně vytvořit společné tělo. Jsou-li však sázeny od sebe dost daleko, svoji přítomnost pocítí až v závěrečné fázi růstu a na přivrácených stranách mohou růst zastavit (obr. 3).
Při zkoumání homospecifického působení dvou morfotypů jsme sázeli kolonie v různých vzdálenostech od sebe. Vzdálenost mezi sousedy v podstatě simuluje stupeň vývoje, při němž se kolonie setkají. Pokud se setkají mladší vývojová stadia, mohou se vzájemně dotknout, eventuálně vytvořit společné tělo. Jsou-li však sázeny od sebe dost daleko, svoji přítomnost pocítí až v závěrečné fázi růstu a na přivrácených stranách mohou růst zastavit (obr. 3).
Dvě kolonie R sázené ve vzdálenosti 10 mm srostou, i když s velmi zachovalou a dobře viditelnou „demarkační“ linií (obr. 3A). Při větších vzdálenostech se na přivrácených stranách růst zastavuje, kolonie k sobě nedorostou a mírně se deformují (do tvaru písmene D). Dvě kolonie F inokulované do vzdálenosti umožňující jejich srůst (3 mm) vytvářejí „soukolonii“ složenou ze dvou středů a společného lemu. Při vzdálenosti 10 mm nesrůstají ani se výrazněji nedeformují, pouze mezi sebou urychleně vytvoří lem (obr. 3B).
Zajímavý je opět pohled na mikroskopickou úroveň interakcí. Pokud dvě kolonie stejného druhu nesrůstají, vysílají na přivrácených stranách předvoj skautujících buněk i poté, co růst zcela zastavily (obr. 3b, d). Naopak na odvrácených stranách volné buňky téměř nevznikají, podobně jako u plně vyvinutých kolonií. Jako by skauti sloužili k zabírání teritoria a přípravě podkladu pro šíření kolonie, k „značkování“ terénu a vymezení se vůči sousedovi, který by se mohl příliš přiblížit. Podobnou situaci pozorujeme i při srůstu dvou stejných kolonií. V místě srůstu jsou opět skauti, kdežto na odvrácených stranách se téměř nevyskytují (obr. 3a, c).
Vzájemné ovlivňování kolonií různých druhů již bylo částečně nastíněno v odstavci o vzniku struktury X. Využili jsme možnosti, kterou skýtá různá ornamentace jednotlivých typů kolonií. Do nestejně starých kolonií Fw (0–24 h) jsme naočkovali biomasu R. Pokud byla přidána k vnějšímu obvodu kolonie Fw, vždy celou kolonii obrostla (obr. 4B), pokud do jejího vnitřního okraje, forma R vycestovala a opět obrostla celou kolonii Fw, ovšem jen tehdy, byla-li kolonie Fw mladší než 24 hodin. Pokud byla biomasa R přidána později, R si cestu ven neprorazil a zůstal uvnitř kolonie Fw uvězněn okolním nárůstem fontánkové kolonie.
Vzájemné interakce druhů S. marcescens (F) a S. rubidaea (R) byly natolik překvapivé, že jsme se rozhodli přidat další bakteriální druh, a to jeden z laboratorních, nepříliš fotogenických kmenů Escherichia coli (obr. 1C vlevo). Fontánková kolonie se v přítomnosti E. coli nikdy nenechá obrůst, naopak v této dvojici přebírá dominantní roli a výrazně brání růstu E. coli směrem k sobě (obr. 2A). Tento zaháněcí trend lze pozorovat dokonce i v případě, že E. coli dostane „náskok“ a biomasa F je k ní přidána až po jednom či dvou dnech samostatného vývoje. Jako „obranná“ reakce na cizí bakteriální tělo opět vzniká výrazná struktura X. Je tím výraznější, čím je větší nárůst sousední E. coli.
Při vzájemném působení mezi koloniemi R a E. coli se situace obrací ve prospěch E. coli. Při současné inokulaci ve vzdálenosti umožňující srůst kolonií (5 mm) E. coli slabě přerůstá R. Pokud však umožníme E. coli alespoň jednodenní náskok, přeroste R úplně. Ve vzdálenosti 15 mm je růst na přivrácených stranách inhibován a mezi koloniemi zůstává volný prostor.
Shrneme-li výsledky heterospecifických interakci, lze konstatovat, že S. rubidaea (R) obrůstá S. marcescens (F) a brání jí v růstu, zároveň S. marcescens (F) omezuje a zahání E. coli a konečně E. coli přerůstá, případně inhibuje S. rubidaea (R). Vzájemné vztahy podřízenosti a nadřazenosti zúčastněných partnerů gradují při nejtěsnějším kontaktu, který vznikne nakapáním směsi dvou klonů v poměru 1 : 1 na malou plochu (obr. 5).
Chiméry
 Směsné kolonie klonů F/Fw a R/W představují setkání stejně silných partnerů. V prvním případě je výsledkem kolonie fontánkového vzhledu, v níž jsou jednotlivé typy buněk přítomny ve středu, mezikruží i lemu (obr. 5A). V druhém případě se výsledný chimerický útvar skládá z kropenatého středu obsahujícího buňky R i W a lemu, v němž se střídají ostře ohraničené paprsky složené z buněk R nebo W (obr. 5B). Ve zbývajících případech (u chimérických útvarů R/E. coli a R/F) určuje výsledný vzhled chiméry dominantní partner. V chiméře R/E. coli je kolem středu složeného ze směsi buněk R a E. coli mohutný lem, který je tvořen výhradně buňkami E. coli (obr. 5C). Podobně je tomu i v chiméře R/F. Směs buněk R a F umístěná ve středu je obklopena buňkami klonu R (obr. 5D). Ani zvětšování inokulačních terčíků základní uspořádání chiméry nemění.
Směsné kolonie klonů F/Fw a R/W představují setkání stejně silných partnerů. V prvním případě je výsledkem kolonie fontánkového vzhledu, v níž jsou jednotlivé typy buněk přítomny ve středu, mezikruží i lemu (obr. 5A). V druhém případě se výsledný chimerický útvar skládá z kropenatého středu obsahujícího buňky R i W a lemu, v němž se střídají ostře ohraničené paprsky složené z buněk R nebo W (obr. 5B). Ve zbývajících případech (u chimérických útvarů R/E. coli a R/F) určuje výsledný vzhled chiméry dominantní partner. V chiméře R/E. coli je kolem středu složeného ze směsi buněk R a E. coli mohutný lem, který je tvořen výhradně buňkami E. coli (obr. 5C). Podobně je tomu i v chiméře R/F. Směs buněk R a F umístěná ve středu je obklopena buňkami klonu R (obr. 5D). Ani zvětšování inokulačních terčíků základní uspořádání chiméry nemění.
Systém vzájemné nadřazenosti ve výsledných vztazích tří sledovaných partnerů S. rubidaea (R), S. marcescens (F) a E. coli se uzavírá do kruhu odpovídajícího známé hře „kámen, nůžky, papír“, kde kamenem je S. rubidaea, nůžkami S. marcescens a papírem E. coli. Celá předchozí pozorování lze shrnout do jednoho malého pokusu. Do bezprostřední blízkosti chiméry složené z morfotypu S. rubidaea a E. coli (R/E. coli) naočkujeme S. marcescens (F). Co pozorujeme? Morfotyp F zastavuje růst E. coli, morfotyp R se proto může vymanit z jejího vlivu, stává se aktivním a pro sousední morfotyp F dominantním, takže fontánku rychle obroste (obr. 6). Abychom však mohli prohlédnout tuto zákulisní hru zúčastněných partnerů, museli jsme projít všemi předchozími pokusy. Úžasným pomocníkem při sledování interakcí nám je bohatá ornamentální variabilita druhů rodu Serratia, ať už jde o barvu, či architekturu kolonií.
Námi popisovaný model dovoluje odhalit a dále studovat vzájemné vztahy několika málo bakteriálních klonů v ontogenezi kolonií. Jednoduchost tohoto komunikačního systému pomůže odhalit „řeč“ vzájemného ovlivňování a prosazování v prostoru a při stavbě kolonie.
Literatura
Čepl J. J., Pátková I., Blahůšková A., Cvrčková F., Markoš A.: Patterning of mutually interacting bacterial bodies: close contacts and airborne signals, BMC Microbiol. 10, 139, 2010.
Rieger T., Neubauer Z., Blahůšková A., Cvrčková F., Markoš A.: Bacterial body plans: colony ontogeny in Serratia marcescens, Communicative Integrative Biology 1, 78–87, 2008.
Věnováno prof. Zdeňku Neubauerovi k významnému životnímu jubileu. Ke gratulaci se připojuje celá laboratoř experimentální filosofie Katedry filosofie a dějin přírodních věd Přírodovědecké fakulty UK: Anna Blahůšková, Anton Markoš, Jaroslav Čepl, Michal Schmoranz a Tomáš Rieger.


 Baktérie, se kterou pracujeme, vytváří obvykle nápadné tmavočervené kolonie se zeleně kovovým leskem. Její platné taxonomické označení Serratia marcescens zní nespravedlivě odpudivě (lat. marcesco = vadnu, hniji). Tradičně je však známa pod přiléhavějším názvem Bacterium prodigiosum („zázračná hůlka“). Za toto označení vděčí tomu, že její červené kolonie prý byly příčinou „krůpějí krve“ na hostiích přechovávaných v uzavřených svatostáncích vlhkých chrámů.
Baktérie, se kterou pracujeme, vytváří obvykle nápadné tmavočervené kolonie se zeleně kovovým leskem. Její platné taxonomické označení Serratia marcescens zní nespravedlivě odpudivě (lat. marcesco = vadnu, hniji). Tradičně je však známa pod přiléhavějším názvem Bacterium prodigiosum („zázračná hůlka“). Za toto označení vděčí tomu, že její červené kolonie prý byly příčinou „krůpějí krve“ na hostiích přechovávaných v uzavřených svatostáncích vlhkých chrámů.
 Naše zkušenosti plně potvrzují přiléhavost tradičního názvu tohoto mikroba, byť ze zcela jiných důvodů, než z jakých mu je připsala pověst. Serratia nevytváří totiž pouze kolonie podobné krvavým krůpějím, ale i řadu dalších. Příčinou je vysoká specifická proměnlivost dědičnosti vzhledu kolonie. Lze ji snadno a s nemalým estetickým požitkem sledovat.
Naše zkušenosti plně potvrzují přiléhavost tradičního názvu tohoto mikroba, byť ze zcela jiných důvodů, než z jakých mu je připsala pověst. Serratia nevytváří totiž pouze kolonie podobné krvavým krůpějím, ale i řadu dalších. Příčinou je vysoká specifická proměnlivost dědičnosti vzhledu kolonie. Lze ji snadno a s nemalým estetickým požitkem sledovat.
Opakované klonování zrajících a stárnoucích populací druhu Serratia marcescens nám umožnilo odhalit skryté zdroje variability této baktérie. V kráse a ve vynalézavosti „jak vypadat“ překonala naše „zázračná baktérie“ všechna očekávání.
Oba citáty jsou z článku Neubauer Z., Koutecká E., Krekulová A.: Proměnlivost a dědičnost vzhledu bakteriálních kolonií, Vesmír 61, 151, 1982/5
Ke stažení
 článek ve formátu pdf [1,44 MB]
článek ve formátu pdf [1,44 MB]
O autorovi
Irena Pátková
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
