









Stále záhadné deinotériá
| 6. 4. 2010Rôzne nejasnosti a dohady sprevádzali deinotériá už od čias ich prvých nálezov v období 18. a 19. storočia. Vášnivú diskusia o systematickej príslušnosti a spôsobe života týchto cicavcov vyvolával najmä nezvyčajný spôsob zakrivenia symfýzy sánky so spodnými klami a svojráznymi zubami impozantných rozmerov. Napriek tomu, že dnešné poznatky o deinotériách sú oveľa kompletnejšie, niektoré otázky sa nepodarilo spoľahlivo zodpovedať ani po 150-ročnom výskume. Ide predovšetkým o tvar a dĺžku chobota a o funkciu spodných klov.
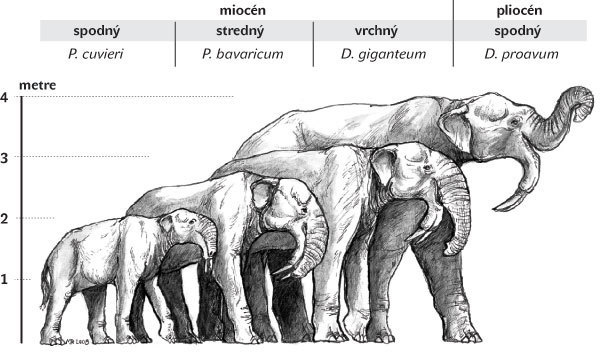
 Deinotériá (Deinotheriidae) reprezentujú najkonzervatívnejšiu, no zároveň vysoko špecializovanú evolučnú líniu archaických chobotnatcov. Predstavujú sesterskú taxonomickú skupinu k slonotvarým (Elephantiformes), ktorá zahŕňa všetky „mastodonty“ a slonovité chobotnatce (mamuty a slony). Aj keď sa v poslednom období boli zaznamenané nové objavy na území Afriky, ktoré posunuli spodnú hranicu stratigrafického rozšírenia deinotérií, o ich pôvode a potenciálnych predkoch nemáme takmer žiadne poznatky. Objavili sa vo vrchnom oligocéne (pred 27–28 mil. r.) a bez výrazných morfologických zmien a fyletických vetvení prežili až do spodného pleistocénu (pred 1,0 mil. r.). Jediným nápadným evolučným trendom deinotérií bolo zväčšovanie telesných rozmerov (obr. 3). Okrem Afriky boli rozšírené takmer na celom území Európy (okrem Škandinávie) a južnej i juhovýchodnej Ázie. Ich migračná schopnosť bola pravdepodobne limitovaná ekologickými faktormi a možnou biotickou konkurenciou „mastodontov“, preto sa nikdy nedostali na územie Ameriky.
Deinotériá (Deinotheriidae) reprezentujú najkonzervatívnejšiu, no zároveň vysoko špecializovanú evolučnú líniu archaických chobotnatcov. Predstavujú sesterskú taxonomickú skupinu k slonotvarým (Elephantiformes), ktorá zahŕňa všetky „mastodonty“ a slonovité chobotnatce (mamuty a slony). Aj keď sa v poslednom období boli zaznamenané nové objavy na území Afriky, ktoré posunuli spodnú hranicu stratigrafického rozšírenia deinotérií, o ich pôvode a potenciálnych predkoch nemáme takmer žiadne poznatky. Objavili sa vo vrchnom oligocéne (pred 27–28 mil. r.) a bez výrazných morfologických zmien a fyletických vetvení prežili až do spodného pleistocénu (pred 1,0 mil. r.). Jediným nápadným evolučným trendom deinotérií bolo zväčšovanie telesných rozmerov (obr. 3). Okrem Afriky boli rozšírené takmer na celom území Európy (okrem Škandinávie) a južnej i juhovýchodnej Ázie. Ich migračná schopnosť bola pravdepodobne limitovaná ekologickými faktormi a možnou biotickou konkurenciou „mastodontov“, preto sa nikdy nedostali na územie Ameriky.
S chobotom, či bez neho?
 Spočiatku boli pre vedu známe len brachyodontné, bunolofodontné (nízkokorunkové hrboľovité premoláre a jarmové moláre) zuby a sánka, čo viedlo k nejednotným a často protikladným hypotézam o presnej identifikácii a systematickom zaradení živočíchov, ktorým tieto nájdené zvyšky prislúchali. Zuby s podobnou morfológiou sa vyskytujú u zástupcov niekoľkých taxonomicky nepríbuzných skupín cicavcov, ktoré sa živia hlavne mäkšou, šťavnatou rastlinnou potravou. Práve vďaka tomuto faktu vznikali nezhody medzi jednotlivými bádateľmi. Fosílie boli nerozhodne priradené k nosorožcom, obrovským tapírom, hrochom, chobotnatcom, sirénam a iným bylinožravým veľrybovitým cicavcom, neraz aj k ich prechodným formám. Najkurióznejší názor predstavoval ich začlenenie medzi vačkovce a leňochy. Nemenej zvláštne boli aj predstavy o používaní spodných klov – sirénam mali slúžiť ako kotva počas spánku na brehoch riek, leňochom na zavesenie na konáre stromov.
Spočiatku boli pre vedu známe len brachyodontné, bunolofodontné (nízkokorunkové hrboľovité premoláre a jarmové moláre) zuby a sánka, čo viedlo k nejednotným a často protikladným hypotézam o presnej identifikácii a systematickom zaradení živočíchov, ktorým tieto nájdené zvyšky prislúchali. Zuby s podobnou morfológiou sa vyskytujú u zástupcov niekoľkých taxonomicky nepríbuzných skupín cicavcov, ktoré sa živia hlavne mäkšou, šťavnatou rastlinnou potravou. Práve vďaka tomuto faktu vznikali nezhody medzi jednotlivými bádateľmi. Fosílie boli nerozhodne priradené k nosorožcom, obrovským tapírom, hrochom, chobotnatcom, sirénam a iným bylinožravým veľrybovitým cicavcom, neraz aj k ich prechodným formám. Najkurióznejší názor predstavoval ich začlenenie medzi vačkovce a leňochy. Nemenej zvláštne boli aj predstavy o používaní spodných klov – sirénam mali slúžiť ako kotva počas spánku na brehoch riek, leňochom na zavesenie na konáre stromov.
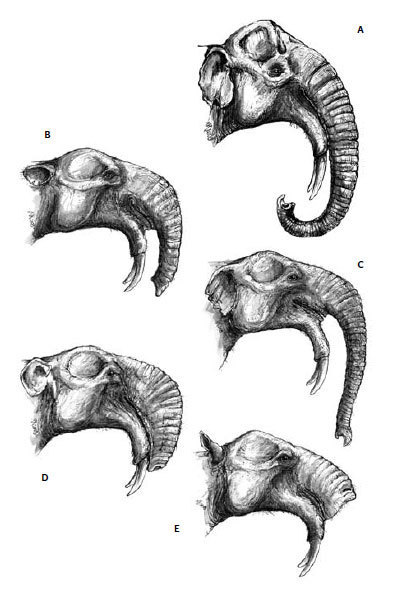
Kľúčové rozlúštenie uvedených zahmlených domnienok a správne nasmerovanie výskumu deinotérií priniesli nálezy takmer kompletných kostier z Opatova (1853) a Františkových Lázní (1883), na základe ktorých boli deinotériá definitívne priradené k chobotnatcom (Augusta, 1938). Následne sa objavili prvé pokusy o „vierohodnejšie“ rekonštrukcie deinotérií, ktoré však neboli ničím iným ako zobrazením „pravých slonov“ so spodnými klami (obr. 2A).
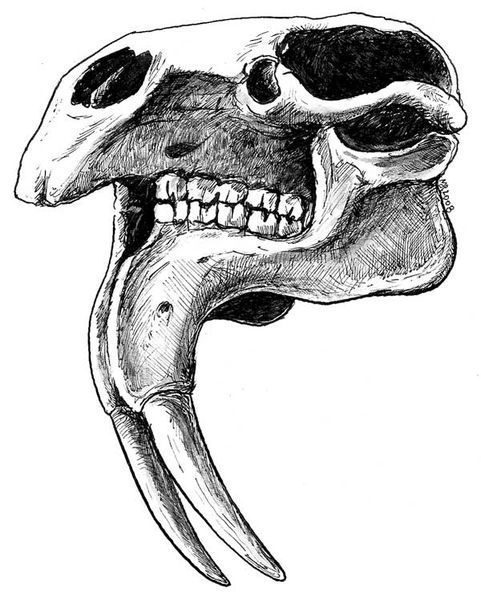
Na začiatku 20. storočia bol tento spôsob zobrazovania spochybnený. Zdokonalením anatomických poznatkov vznikli oveľa reálnejšie predstavy z hľadiska morfológie hlavy, naďalej však zostala otáznou dĺžka a tvar chobota (obr. 2 B,C) (Osborn, 1910; Abel, 1922). Rozmery nosových otvorov, ich nápadné posunutie na lebke smerom dozadu, výrazná prítomnosť V. a VII. faciálneho (trigeminálneho) nervu sú jednoznačné argumenty v prospech existencie chobota. Na rozdiel od zástupcov čeľade slonovitých (Elephantidae), deinotériá mali diametrálne odlišné, z vrchnej strany sploštené a viac-menej horizontálne orientované premaxily, ktoré neposkytovali dostatočný priestor na upnutie svalovej hmoty, aký vyžaduje dĺžka a tvar chobota recentných slonov (obr. 1). Na základe týchto poznatkov sú oveľa reálnejšie predstavy Engessera a kol. (1996) a Markova a kol. (2001), ktorí deinotériá znázorňujú s oveľa kratším chobotom, i keď na jeho tvar majú odlišný názor. Prví menovaní autori zobrazujú deinotériá s mierne predĺženým, splošteným chobotom, tvarovo podobným dnešným tapírom (obr 2B). Podľa Markova a kol. (2001) bol chobot deinotérií svalnatý a mierne rozšírený do strán, pričom sčasti prekrýval spodnú ceľusť (obr. 2D). Treba podotknúť, že počas pomerne dlhej evolučnej histórie deinotérií existoval málo výrazný trend v modifikácii nosovej oblasti lebky, ktorý napovedá o postupnom zväčšovaní chobota. Tento orgán však zjavne nemohol dosahovať veľkosť charakteristickú pre súčasné slony. Otázka, či bol koniec chobota deinotérií ukončený prstovitým výbežkom (resp. výbežkami), schopným komplexného pohybu a manipulácie s potravou, alebo bol morfologicky menej sofistikovaný (podobný „chobotu“ tapírov), zostáva v dôsledku absencie testovateľných dôkazov nezodpovedaná a v čisto hypotetickej rovine.
 Medzi kontroverzné a z hľadiska vedeckej exaktnosti kriticky odmietané patria rekonštrukcie Gregora a kol. (2000), zobrazujúce deinotériá bez chobota, s hranatou hornou perou (obr. 2E). Vychádzali z absencie horných a smerovania spodných klov, čo umožňovalo priamy kontakt ústneho otvoru s potravou, bez nevyhnutnej pomoci chobota. Autori však neakceptovali spomínané morfologické znaky lebky. Takýto vzhľad je nereálny aj z hľadiska problematického fyzického kontaktu hlavy so zemským povrchom, ktorý je nevyhnutný napríklad pri pití vody. Naznačený problém mohol byť eliminovaný len v prípade, ak by deinotériá preferovali močaristé alebo iné vodné biotopy. Proti takémuto spôsobu života však hovoria anatomické proporcie končatín, ktoré jednoznačne indikujú adaptáciu na pohyb po tvrdšom teréne.
Medzi kontroverzné a z hľadiska vedeckej exaktnosti kriticky odmietané patria rekonštrukcie Gregora a kol. (2000), zobrazujúce deinotériá bez chobota, s hranatou hornou perou (obr. 2E). Vychádzali z absencie horných a smerovania spodných klov, čo umožňovalo priamy kontakt ústneho otvoru s potravou, bez nevyhnutnej pomoci chobota. Autori však neakceptovali spomínané morfologické znaky lebky. Takýto vzhľad je nereálny aj z hľadiska problematického fyzického kontaktu hlavy so zemským povrchom, ktorý je nevyhnutný napríklad pri pití vody. Naznačený problém mohol byť eliminovaný len v prípade, ak by deinotériá preferovali močaristé alebo iné vodné biotopy. Proti takémuto spôsobu života však hovoria anatomické proporcie končatín, ktoré jednoznačne indikujú adaptáciu na pohyb po tvrdšom teréne.
Kly na hrabanie?
Nedoriešená je aj funkcia spodných klov. Zvláštne tvarované a orientované kly (homologické s druhými spodnými rezákmi) sú prítomné počas celej evolúcie deinotérií takmer v nezmenenej forme aj veľkosti. To nasvedčuje, že kly rozhodne museli mať nejakú úlohu. Špeciálna morfológia tylových hrboľov lebky ako aj opačné zakrivenie výbežkov (zygapofýz) predných krčných stavcov deinotérií v porovnaní s dnešnými slonmi umožňovali väčšiu pohyblivosť krku a hlavy do strán aj vertikálnym smerom, čo mohlo rozšíriť aj možnosti využitia klov. Ich osobité smerovanie v kombinácii s celkovou telesnou stavbou však limituje takmer všetky druhy činnosti, ktoré mohli potenciálne vykonávať. Existujú názory, že kly mali iba signalizačnú (imponačnú), resp. obrannú funkciu. Z dôvodu málopočetného a fragmentárneho charakteru fosílneho materiálu dosiaľ nebol preukázaný pohlavný dimorfizmus na základe rozmerov a morfológie klov, čo však neznamená, že neexistoval. Teoreticky tým však nemožno vylúčiť význam klov v sociálnom živote deinotérií.
Masívna symfýza „vyzbrojená“ mohutnými klami vizuálne pôsobí ako impozantný obranný nástroj, ale kly mohli byť použiteľné iba v bezprostrednej blízkosti nepriateľa. Preto takmer s istotou môžeme vylúčiť ich aplikáciu na obrannú činnosť. Navyše už samotné telesné rozmery deinotérií mohli poskytnúť adekvátnu ochranu pred akýmkoľvek predátorom. O aktívnom používaní klov hovorí fakt, že juvenilné jedince mali kly dlátovito zakončené, s vrstvou skloviny. Až neskôr nadobudli mechanickým obrúsením typický kónický tvar, pričom sklovina bola odstránená a hmotu klov tvoril len masívny dentín.
Napriek tomu kly niektorých úplne dospelých jedincov vykazujú len menšie stopy po mechanickom opotrebovaní. To nasvedčuje, že používanie klov bolo individuálne, značne premenlivé a závislé aj od lokálnych podmienok prostredia. S obdobným javom sa stretávame aj u recentných slonov.
Najreálnejšou úlohou klov mohla byť pomoc pri získavaní potravy. Rozšírenou predstavou je, že slúžili na vyhrabávanie podzemných častí rastlín. Celková telesná stavba, rozmery, ako aj anatomická pozícia samotných klov (pomerne veľká vzdialenosť od ich konca k zemskému povrchu) nasvedčujú, že takúto činnosť mohli vykonávať len vo veľmi neprirodzenej polohe (kľačiačky) alebo na vyvýšenom mieste nad úrovňou terénu, kde zviera stálo (napr. na brehoch riek). Obidve situácie sú však málo pravdepodobné. Okrem toho pri hrabaní by sa do ústneho otvoru spolu s rastlinnými zvyškami dostalo aj množstvo pôdnych zŕn, čo by počas evolučného procesu určite viedlo k hypsodoncii (nárast výšky korunky) zubov. Táto však vôbec nie je pozorovaná. (Harris, 1975).
Anatomické modifikácie hlavy a krčných stavcov umožňovali pomerne širokú škálu pohybov, čo bolo výhodné pri spásaní šťavnatej vegetácie, pravdepodobne však len z vyšších vegetačných poschodí lesných biotopov. Takúto adaptačnú stratégiu naznačuje aj konzervatívna morfológia zubov, relatívne krátke kly a chobot, ako aj celkové telesné rozmery deinotérií. V procese zaobstarávania potravy mohli kly slúžiť na selektívne získavanie určitých častí rastlín, napr. lámanie vetvičiek drevín alebo sťahovanie vysoko výživnej kôry stromov. Dokladajú to aj zbrúsené plochy hlavne na prednej strane klov (z hľadiska anatomickej pozície ide v skutočnosti o zadnú stranu týchto zubov). V prípade takejto aplikácie klov by sa však potrava sústreďovala v blízkosti predných končatín a na jej dosiahnutie by bol potrebný dlhý chobot, ktorý by však pri činnosti klov určite prekážal a musel by byť stočený vo zvláštnej anatomickej polohe. Z tohto hľadiska je reálnejší a výhodnejší kratší chobot. Kly nemuseli zohrávať hlavnú úlohu pri zabezpečovaní potravy, ale mohli sa používať len príležitostne, na špeciálne úkony, pričom primárnu funkciu získavania potravy mohol spĺňať chobot.
Z uvedených poznatkov vyplýva, že prakticky s istotou vieme eliminovať dĺžku, ktorú chobot deinotérií nemohol dosahovať, resp. aké funkcie spodné kly nedokázali vykonávať. S určitosťou môžeme prehlásiť, že chobot deinotérií bol v porovnaní s dnešnými slonmi kratší, jeho tvar však zostáva záhadou. Či bola funkcia klov obmedzená na jedinú, alebo mohli vykonávať viacero odlišných činností, zostáva z dôvodu unikátnosti týchto elementov a absencie analogických prípadov z recentnej živočíšnej ríše naďalej otvorenou otázkou.
Literatura
Abel O.: Lebensbilder aus der Tierwelt der Vorzeit, Jena, 1922
Augusta J.: Stará zpráva o nálezu Dinotheria u Opatova, Čas. Nár. musea v Praze, odd. přírod., 112, 32–48, 1938
Engesser B., Fejfar O., Major P.: Das Mammut und seine ausgestorbenen Verwandten, Veröffentlichungen aus dem Naturhistorischen Museum Basel 20, 1–188, 1996
Gregor H.-J., Kuhn R., Storch D. H.: Deinotherium? Ein Proboscidier? Gedanken zur taxonomisch-systematischen Stellung von Deinotherium auf Grund anatomisch-morphologischer Gegebenheiten, Documenta Naturae (Munich) No. 130, 1–142, 2000
Harris J. M.: Evolution of feeding mechanism in the family Deinotheriidae (Mammalia, Proboscidea), Zool. J. Linn. Soc. 56, 331–362, 1975.
Markov G. N., Spassov N., Simeonovski V.: A Reconstruction of the Facial Morphology and Feeding Behavior of the Deinotheres. The World of Elephants, International Congress, Rome 2001
Osborn H. F.: The Age of Mammals in Europe, Asia and North America, Mac- Millan, New York 1910
Slovníček
Sánka – dolná čeľusť (lat. mandibula)
symfýza sanky – predná časť sánky, kde sa spája jej pravá a ľavá horizontálna vetva. U deinotérií zo symfýzy vyrastajú spodné kly.
premolár – (lat. dens premolaris) čreňový zub cicavcov
molár – (lat. dens molaris) stolička cicavcov
Ke stažení
 článek ve formátu pdf [294,42 kB]
článek ve formátu pdf [294,42 kB]
O autorovi
Csaba Tóth
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
