










Rychlejší vyhrává
 Zelené rostliny nezbytně potřebují světlo a to se k nim dostává různými cestami. Pohyb listů ve větru může rostliny stínit v sekundových intervalech, pohyb oblaků v intervalech minutových či hodinových. Přitom rozdíl mezi intenzitou světla v přímém slunečním světle a v hlubokém stínu v zápoji lesní vegetace může být až tři řády. Na spodních listech proto kolísá světlo od hodnot, kdy fotosyntéza sotva stačí vyrovnávat dýchání a rostlina neroste, po plný sluneční svit, který nepřipravený list poškozuje. A právě fotosyntetické připravenosti listu si všimneme blíže.
Zelené rostliny nezbytně potřebují světlo a to se k nim dostává různými cestami. Pohyb listů ve větru může rostliny stínit v sekundových intervalech, pohyb oblaků v intervalech minutových či hodinových. Přitom rozdíl mezi intenzitou světla v přímém slunečním světle a v hlubokém stínu v zápoji lesní vegetace může být až tři řády. Na spodních listech proto kolísá světlo od hodnot, kdy fotosyntéza sotva stačí vyrovnávat dýchání a rostlina neroste, po plný sluneční svit, který nepřipravený list poškozuje. A právě fotosyntetické připravenosti listu si všimneme blíže.
List přenesený ze tmy na sluníčko nezačne hned fotosyntetizovat naplno, trvá to půl hodiny i více. Toho si prvně všimnul již W. J. W. Osterhout v roce 1918. Může za to několik procesů, které se rozbíhají současně, ale nikoli stejně rychle:
- Světelnými reakcemi se regeneruje substrát potřebný pro vázání oxidu uhličitého, ribulóza-1,5-bisfosfát (1–2 minuty).
- Aktivuje se enzym rubisco,1 který váže CO2 na tento substrát (5–10 minut).
- Otevírají se průduchy, které v uzavřeném stavu brání difuzi CO2 do listu (20 minut i déle).
Pro rostliny otevřených stanovišť neznamená ztráta času při otevírání průduchů velký problém. Slunce na ně svítí skoro celý den. Fotosyntéza ráno naběhne a není-li omezována například suchem nebo zamračenou oblohou, běží s plnou rychlostí až do večera. Zato v podrostu tropických deštných lesů je po většinu dne „fotosyntetická tma“, která je prozařována jen prchavými světelnými skvrnami. 2) Přestože trvání všech prchavých světýlek dohromady je proti trvání hlubokého stínu často zanedbatelné, jsou až tisíckrát intenzivnějším zdrojem světelné energie než nepřímé světlo pronikající přes listovou mozaiku. Přinášejí tak 30–90 % z celkového množství světelných kvant dopadajících na lesní podrost. Rostlina v podrostu je na využívání energie těchto záblesků světla závislá, a proto i drobné zdokonalení v rychlosti „nabíhání fotosyntézy“ pro ni může znamenat velkou výhodu.
Bylo napsáno mnoho prací o tom, jak podrostové druhy a semenáčky využívají drobné a pomíjivé flíčky slunečního světla. Na jednu stranu se ukazuje, že rostliny vyrůstající v kolísavém světle jsou přizpůsobeny tak, aby aktivaci fotosyntézy urychlovaly, a naopak si tuto aktivovanost déle udržely i ve stínu a byly připraveny na další světelný flíček. Na stranu druhou je zjevné, že jejich znevýhodnění je zčásti „vrozené“ a zdá se být nepřekonatelné. Evoluce, jak známo, nepracuje systematicky. To, co vytvořila, je dílem náhody a zachovalo se to proto, že to ještě v celistvosti nebylo překonáno něčím úspěšnějším.
V tomto okamžiku se přesto nabízí otázka, proč zmíněné tři procesy neprobíhají rychleji, ačkoli by to bylo evolučně výhodné. Odpovědi na ni jsou v současnosti bohužel spíše spekulativní. Přesto se u těchto spekulací trochu pozastavíme.
 První (nejrychlejší) proces závisí na energii potřebné pro vytvoření substrátu, na který se naváže CO2. Z množství substrátu vyplývá energetický požadavek, a tedy čas potřebný k aktivaci. Toto množství je většinou takové, že vystačí na pokračování fotosyntézy dalších 10 až 20 sekund po zhasnutí. Vidíme tedy, že vytváření zásob trvá déle, než na kolik následná fotosyntéza vystačí. Tato asymetrie je zajímavá, ale nikoli nevysvětlitelná. Souvisí s dalšími dvěma faktory, které se teprve aktivují. Po jejich pochopení se nám logický kruh uzavře.
První (nejrychlejší) proces závisí na energii potřebné pro vytvoření substrátu, na který se naváže CO2. Z množství substrátu vyplývá energetický požadavek, a tedy čas potřebný k aktivaci. Toto množství je většinou takové, že vystačí na pokračování fotosyntézy dalších 10 až 20 sekund po zhasnutí. Vidíme tedy, že vytváření zásob trvá déle, než na kolik následná fotosyntéza vystačí. Tato asymetrie je zajímavá, ale nikoli nevysvětlitelná. Souvisí s dalšími dvěma faktory, které se teprve aktivují. Po jejich pochopení se nám logický kruh uzavře.
Druhým, složitějším a variabilnějším faktorem je otevírání a zavírání průduchů. Průduchy jsou pro přežívání suchozemských rostlin klíčové, protože umožňují regulovat výpar vody z listu (transpiraci). Zároveň s omezením ztráty vody se však omezí i příjem CO2. Většina rostlin zavírá průduchy nejen při nedostatku vody, ale také při nedostatku světla. Logiku to má. Bez světla není fotosyntéza, tak proč zbytečně ztrácet vodu… A právě v tom je ten problém! Světlo může přijít velmi rychle, ale průduchy se otvírají 20 až 90 minut 3) a u většiny rostlin pulzují i při dostatku vody. Existují sice tropické podrostové druhy, které ve stínu průduchy nezavírají úplně, nechávají je pootevřené. V tropickém pralese bývá při zemi téměř 100% vlhkost vzduchu, což znamená nulovou transpiraci. Nadměrný výpar zde rostlinám věru nehrozí. Proč tedy nenechají průduchy „dokořán“, aby byly připraveny na krátkodobý záblesk slunce? Nevíme. Přitom pomalé pulzování průduchů (právě proto, že je dlouhotrvající) způsobuje největší ztrátu potenciálně získatelného CO2, a tedy nenarostlé biomasy rostliny.
 Zajímavá je jedna adaptace, která staví průduchy během fotosyntetické indukce „mimo hru“. Nedávno se zjistilo, že se v tropickém podrostu vyskytují rostliny, které otevírají průduchy na noc a vážou CO2 do organických kyselin. Ve dne jej odbouráním skupiny COOH (již se zavřenými průduchy) uvolňují a pomocí slunečního světla definitivně vážou do organických látek. Tak to dělají mnohé sukulenty. U kaktusů v poušti to znamená značnou úsporu vody (v noci je tam zima a vlhko, takže výpar otevřenými průduchy je minimální). U podrostových druhů, např. ananasovníku (Ananas comosus) či bromélie (Bromelia plumieri), se zase využije vysoká koncentrace CO2 v listu způsobená uvolňováním CO2 z kyselin při zavřených průduších. Hned se ovšem nabízí otázka, proč tedy takto přizpůsobené rostliny v tropickém podrostu nepřevládají. Může to být tím, že jejich energetický metabolizmus je méně účinný (potřebuje větší množství světla na stejné množství získaného CO2).
Zajímavá je jedna adaptace, která staví průduchy během fotosyntetické indukce „mimo hru“. Nedávno se zjistilo, že se v tropickém podrostu vyskytují rostliny, které otevírají průduchy na noc a vážou CO2 do organických kyselin. Ve dne jej odbouráním skupiny COOH (již se zavřenými průduchy) uvolňují a pomocí slunečního světla definitivně vážou do organických látek. Tak to dělají mnohé sukulenty. U kaktusů v poušti to znamená značnou úsporu vody (v noci je tam zima a vlhko, takže výpar otevřenými průduchy je minimální). U podrostových druhů, např. ananasovníku (Ananas comosus) či bromélie (Bromelia plumieri), se zase využije vysoká koncentrace CO2 v listu způsobená uvolňováním CO2 z kyselin při zavřených průduších. Hned se ovšem nabízí otázka, proč tedy takto přizpůsobené rostliny v tropickém podrostu nepřevládají. Může to být tím, že jejich energetický metabolizmus je méně účinný (potřebuje větší množství světla na stejné množství získaného CO2).
 Ještě větší záhadou je role enzymu rubisco. Jeho regulace světlem je sice potřebná k ochraně před biochemickými nerovnováhami, ale časový horizont už tak samozřejmý není. Protože mechanizmus aktivace rubisco je poměrně složitý, je pravděpodobné, že jde o evoluční pozůstatek. Rubisco je vůbec velmi nemoderní enzym. Jeho molekula je velká a katalytická rychlost velmi pomalá (proto ho v rostlinách musí být takové množství, že je považován za nejrozšířenější bílkovinu na Zemi), a navíc si „plete“ oxid uhličitý s kyslíkem. Proč tedy nevěřit, že i jeho pomalá aktivace je pozůstatkem ze starých časů, kdy bylo ve vzduchu mnohem více CO2, a tak to tolik nevadilo? Období, kdy je obsah oxidu uhličitého ve vzduchu menší než řekněme 0,1 %, je evolučně poměrně krátké, přibližně posledních 5 milionů let. Vidíme, že zmíněné tři složky fotosyntetické indukce spolu úzce souvisejí a dohromady způsobují velké znevýhodnění podrostových rostlin. Uzavírá se nám tak slibovaný logický kruh – díky nízké počáteční aktivitě rubisco a zavřeným průduchům je pomalá i počáteční syntéza substrátu pro vázání CO2, která trvá mnohem déle (1–2 minuty) než pokračující plná fotosyntéza po zhasnutí (10 až 20 sekund).
Ještě větší záhadou je role enzymu rubisco. Jeho regulace světlem je sice potřebná k ochraně před biochemickými nerovnováhami, ale časový horizont už tak samozřejmý není. Protože mechanizmus aktivace rubisco je poměrně složitý, je pravděpodobné, že jde o evoluční pozůstatek. Rubisco je vůbec velmi nemoderní enzym. Jeho molekula je velká a katalytická rychlost velmi pomalá (proto ho v rostlinách musí být takové množství, že je považován za nejrozšířenější bílkovinu na Zemi), a navíc si „plete“ oxid uhličitý s kyslíkem. Proč tedy nevěřit, že i jeho pomalá aktivace je pozůstatkem ze starých časů, kdy bylo ve vzduchu mnohem více CO2, a tak to tolik nevadilo? Období, kdy je obsah oxidu uhličitého ve vzduchu menší než řekněme 0,1 %, je evolučně poměrně krátké, přibližně posledních 5 milionů let. Vidíme, že zmíněné tři složky fotosyntetické indukce spolu úzce souvisejí a dohromady způsobují velké znevýhodnění podrostových rostlin. Uzavírá se nám tak slibovaný logický kruh – díky nízké počáteční aktivitě rubisco a zavřeným průduchům je pomalá i počáteční syntéza substrátu pro vázání CO2, která trvá mnohem déle (1–2 minuty) než pokračující plná fotosyntéza po zhasnutí (10 až 20 sekund).
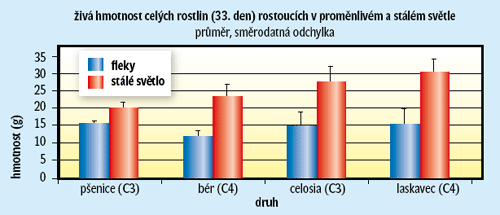
Proměnlivý světelný režim omezuje fotosyntézu a produkci nejen lesních společenstev, ale i společenstev nelesních, polních plodin, a dokonce vodních řas. Pěstovali jsme čtyři bylinné druhy v růstovém boxu jednak při stálém světle, jednak při střídající se silné a slabší ozářenosti. Přitom silná ozářenost byla desetkrát intenzivnější než slabá (v souladu se světelným klimatem v nelesních společenstvech). Světelné skvrny byly různě dlouhé (1–10 minut) a bylo jich během dne asi 100. Důležité je, že světelný program byl nastaven tak, aby jak při stálém světle, tak při světelných skvrnách rostliny obdržely během dne stejné množství světla. Kdyby tedy rostliny dokázaly využívat proměnlivé světlo stejně efektivně jako středně intenzivní stálé, byly by jejich růst a průměrná fotosyntéza v obou režimech vyrovnané. Ve skutečnosti je mezi nimi velký rozdíl (viz obrázek 4). V lesních porostech s hlubšími obdobími stínu bývají někdy extrémy ještě větší. Tady mohou být ve výhodě například mechorosty a lišejníky (obrázek 1) a v nevýhodě naopak rostliny s koncentračním mechanizmem CO2 (obrázek 3). Téma využívání proměnlivého světla, které je v přírodě tak běžné, není tedy aktuální jen pro základní výzkum, ale hlavně pro zemědělce a lesníky, kterým snižuje výnos. A možná brzy zaujme i genetické inženýry, kteří se pokusí vytvořit výnosnější plodiny (nebo i dřeviny) s lepší efektivitou využívání světlených skvrn. Obávám se však, že to bude cesta trnitá (odhlédnuto od otázky, zda je eticky správná). Udělat „lepší“ rubisco usiluje již mnoho let několik pracovních skupin na světě, zatím bez valného úspěchu.
Poznámky
JAK SE MĚŘÍ FOTOSYNTÉZA A PRODUKCE
V ekologii a fyziologii se s potřebou měřit krátkodobou i dlouhodobou rostlinnou produkci (fotosyntézu i hromadění biomasy) setkáváme velmi často. Biomasu vyčíslíme bez složitých a drahých přístrojů. Rostlinu (nebo její sledované části) sklidíme, sušíme při 105 °C tak dlouho, až se hmotnost nemění, a pak hmotnost zapíšeme. Pro měření fotosyntézy potřebujeme složitější techniku, například přístroj pro měření plynové výměny LI-6400 (viz obrázek 2). Princip je jednoduchý. Do komory umístěné v rukojeti se uzavře list, svítí se na něj a měří se koncentrace CO2 a vody před vstupem do komory a po výstupu z ní. Fotosyntetizující list odebírá CO2 a vypařuje vodu. Přístroj měří tyto rozdíly a také průtok vzduchu komorou, což už stačí pro výpočet rychlosti fotosyntézy a transpirace listu. LI-6400 však dokáže měřit a počítat řadu dalších veličin, např. ozářenost, teplotu listu a vzduchu, vodivost průduchů, koncentraci oxidu uhličitého v listu. Je to velmi užitečný a všestranný pomocník fotosyntetického výzkumu. Jeho cena přesahuje v přepočtu na naši měnu 1 milion korun.Ke stažení
 článek v souboru pdf [382,11 kB]
článek v souboru pdf [382,11 kB]
O autorovi
Jiří Kubásek

Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
