










Biologické hodiny u rostlin
| 5. 1. 1996Ve Vesmíru 70, 68, 1991/2 a Vesmíru 73, 425, 1994/8 jsme si mohli přečíst poutavé články H. Illnerové o biologických hodinách v mozku. Neméně fascinující rytmicitu, dokonce s daleko pestřejší fenomenologií, známe u rostlin. Snad každý z nás byl někdy okouzlen pravidelným opakováním ročních cyklů růstu a vývoje rostlin, zachyceným tak přesvědčivě třeba v českých názvech měsíců květen či listopad. Samozřejmě si klademe otázku, jak je tohle pravidelné opakování zajištěno. V našem temperátním klimatu (s cyklem čtyř ročních období) hraje hlavní úlohu střídání světla a tmy a svůj význam mají i změny teploty. To ovšem znamená, že rostliny musí mít systémy rozpoznání, příjmu a zpracování takových vnějších signálů. A nejen to, ukázalo se, že součástí vývojového chování rostlin, jež znamená přizpůsobení sezónnosti počasí, jsou i biologické hodiny projevující se endogenní rytmicitou. Jak tedy vypadá to spojení chronometru nebeského a rostlinného? O vysvětlení se nelze pokusit bez předběžného uvedení poznatků o endogenní rytmicitě rostlin a jejich příjmu světelných signálů. Začneme malou kapitolou z historie. Nejen pro zajímavost, ale hlavně k ozřejmení logiky, kterou se v této souvislosti věda ubírala.
Historie objevu endogenní rytmicity a fotoperiodizmu
Cyklické střídání různých dějů při střídání světla a tmy, např. otevírání a zavírání květů, změny v rychlosti fotosyntézy a další, patří mezi obecně popisované jevy. Některé z nich mohou projevovat cyklický charakter i v nepřetržité temnotě, někdy výjimečně i při stálém osvětlení, tedy v podmínkách, kdy byla zrušena cykličnost působení podnětů (signálů), s nimiž zmíněné jevy spojujeme. Na tuto skutečnost poprvé upozornil ve zprávě Francouzské akademie věd astronom Jean Baptiste de Mairan r. 1729. Předmětem jeho sdělení byl pohyb listů heliotropních (na světlo reagujících) rostlin v trvalé temnotě. Pomocí kimografu registroval jak vzpřimování, k němuž v přírodních podmínkách dochází na světle, tak svěšení, spojované s nástupem temnoty (obrázek). Také následující práce, mezí jejichž autory se vyskytuje i jméno Darwinovo, se týkaly pohybu listů. Zaměřovaly se především na vyloučení dalších možných zdrojů rytmicity, např. změn ve složení ovzduší. Rostliny byly uzavírány do kufrů, vinných sklepů či jeskyň, a nakonec se v plném rozsahu potvrdil de Mairanův fenomén. Okolnost, že byl po dlouhé období studován okrajový jev, znamenala i odsunutí celé problematiky na okraj zájmu fyziologů, byť poučení o možné existenci jakési vnitřní rytmicity rostlin, spojené snad i s měřením času, zůstalo.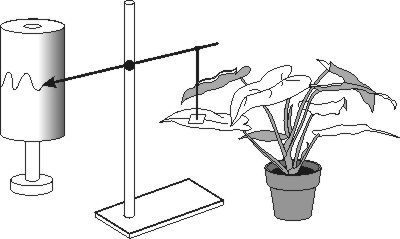 Další krok následoval téměř o dvě stě let později. V ročence Ministerstva zemědělství USA z r. 1920 zobecnili Garner a Allard svá pozorování o vlivu změny délky dne na kvetení rostlin: Zkracující se den vyvolal kvetení sóje a tabáku, zatímco jiné druhy reagovaly iniciací květů na prodlužující se den. Autoři tento jev nazvali fotoperiodizmus a provedli podle typu fotoperiodické reakce kategorizaci rostlin na krátkodenní a dlouhodenní, popřípadě neutrální. Z pozorovaného jevu vyplývá, že existuje mechanizmus velmi citlivého měření délky dne či noci, jež rostlina spojuje s určením ročního období. Další pokrok znamenalo až překvapivé spojení pohybu listů v temnotě s fotoperiodicky vyvolaným kvetením. Takový krok učinil r. 1936 jenský fyziolog Erwin Bünning, který se úspěšně pokusil najít společného jmenovatele pro jevy fotoperiodizmu i endogenní rytmicity, pro objekty rostlinné i živočišné. Souhrn práce byl obsažen již v názvu „Die endonome Tagesrhythmik als Grundlage der photoperiodischen Reaktion“ („Endogenní denní rytmy jako základ fotoperiodické reakce„). Předpokládal, že mechanizmus fotoperiodické regulace kvetení je totožný s mechanizmem odpovědným za pohyby listů v temnotě. Uvažoval časoměrný mechanizmus se dvěma alternujícími fázemi – fotofilní (světlomilnou) a skotofilní (temnomilnou). Výsledek fotoperiodického chování rostliny pak závisí na tom, ve které fázi je světlo přijímáno. Rytmicita, na níž byla celá teorie založena, měla cirkadiánní (dies – den) charakter, tedy její perioda měla délku přibližně jednoho dne (u rostlin byla posléze zjištěna řada endogenních rytmů s periodami kratšími, od přibližně půldenních až k minutovým).
Další krok následoval téměř o dvě stě let později. V ročence Ministerstva zemědělství USA z r. 1920 zobecnili Garner a Allard svá pozorování o vlivu změny délky dne na kvetení rostlin: Zkracující se den vyvolal kvetení sóje a tabáku, zatímco jiné druhy reagovaly iniciací květů na prodlužující se den. Autoři tento jev nazvali fotoperiodizmus a provedli podle typu fotoperiodické reakce kategorizaci rostlin na krátkodenní a dlouhodenní, popřípadě neutrální. Z pozorovaného jevu vyplývá, že existuje mechanizmus velmi citlivého měření délky dne či noci, jež rostlina spojuje s určením ročního období. Další pokrok znamenalo až překvapivé spojení pohybu listů v temnotě s fotoperiodicky vyvolaným kvetením. Takový krok učinil r. 1936 jenský fyziolog Erwin Bünning, který se úspěšně pokusil najít společného jmenovatele pro jevy fotoperiodizmu i endogenní rytmicity, pro objekty rostlinné i živočišné. Souhrn práce byl obsažen již v názvu „Die endonome Tagesrhythmik als Grundlage der photoperiodischen Reaktion“ („Endogenní denní rytmy jako základ fotoperiodické reakce„). Předpokládal, že mechanizmus fotoperiodické regulace kvetení je totožný s mechanizmem odpovědným za pohyby listů v temnotě. Uvažoval časoměrný mechanizmus se dvěma alternujícími fázemi – fotofilní (světlomilnou) a skotofilní (temnomilnou). Výsledek fotoperiodického chování rostliny pak závisí na tom, ve které fázi je světlo přijímáno. Rytmicita, na níž byla celá teorie založena, měla cirkadiánní (dies – den) charakter, tedy její perioda měla délku přibližně jednoho dne (u rostlin byla posléze zjištěna řada endogenních rytmů s periodami kratšími, od přibližně půldenních až k minutovým).
 Zatím poslední kapitolu historie napsali opět fyziologové ve službách amerického Ministerstva zemědělství. Roku 1946 skupina z Beltsvillu identifikovala univerzální receptor fotoperiodického signálu a popsala jeho funkci. Šlo o chromoprotein fytochrom. To jsou tedy milníky na cestě výkladu fotoperiodických jevů vývoje rostlin regulovaných za účasti endogenní rytmicity.
Zatím poslední kapitolu historie napsali opět fyziologové ve službách amerického Ministerstva zemědělství. Roku 1946 skupina z Beltsvillu identifikovala univerzální receptor fotoperiodického signálu a popsala jeho funkci. Šlo o chromoprotein fytochrom. To jsou tedy milníky na cestě výkladu fotoperiodických jevů vývoje rostlin regulovaných za účasti endogenní rytmicity.
Endogenní rytmy rostlin
Tyto rytmy jsou charakterizovány průběhem nezávislým na oscilacích faktorů vnějšího prostředí. Zaměříme se na ty, jež mají cirkadiánní charakter. Předpokládá se existence primárního oscilátoru, pacemakeru, kterým rostliny měří čas. Na rozdíl od vyšších živočichů, u nichž je lokalizace oscilátoru známa (v suprachiazmatickém jádře hypotalamu), nevíme o lokalizaci a funkci rostlinného oscilátoru téměř nic. Není ani jasné, zda existuje jeden univerzální oscilátor, na který jsou nějakým způsobem napojeny všechny rytmické procesy, či zda existuje více oscilátorů specializovaných. U rostlin byla popsána řada cirkadiánních rytmů, počínaje již zmiňovaným pohybem listů, rytmů kvetení (obrázek), růstu řady orgánů a enzymových aktivit. Velmi detailně byl prostudován rytmus metabolizmu CO2 u Bryophyllum fedtschenkoi, důkladně jsou popsány i rytmy u rostlin nižších, např. bioluminiscence u hnědé řasy Gonyaulax polyedra, konidiace u houby Neurospora crassa a buněčné dělení u zelené řasy Euglena gracilis. Ve všech těchto případech však sledujeme pouze pohyb „hodinových ručiček“ a ne pohyb a mechanizmus „stroje“, který je řídí. Významná cirkadiánní rytmicita byla pozorována i v transkripční aktivitě a existují nepřímé důkazy (ovlivnění fáze i periody etanolem či těžkou vodou), poskytující motivace pro hledání oscilátoru v genomu. Ze všech získaných výsledků lze vyvodit, že u rostlin nemá oscilátor specifickou lokalizaci. Při řízení autonomních pohybů listů je zdrojem rytmicity pulvinus („hybný polštářek“) řapíku listu, při udržování rytmu v metabolizmu CO2 u Bryophyllum je autonomní epidermis (pokožka), ne však již mezofyl (vnitřní pletivo) listu. V souladu s hromadícími se důkazy funkce c-AMP (cyklického adenosinmonofosfátu) a inositoltrisfosfátu jako druhých poslů (látek přenášejících informaci z membrány do nitra buňky) se i u rostlin ukazuje, že tyto látky se mohou významně podílet v regulaci rytmicity. Vzhledem k tomu, že inositolový signální systém je úzce propojen s regulací hladiny vápníku v buňce, nepřekvapí nás, že i jeho hladina může ovlivňovat rytmicitu. Ostatně jak se ukázalo při studiu pohybu listů, i změny toků dalších iontů (protonů, draslíku, chloridových) mohou mít pro rytmicitu význam. Potíž je v tom, že i na úrovni druhých poslů a změn transkripce jsme stále blíže ručičkám než hodinovému stroji.
Ze všech získaných výsledků lze vyvodit, že u rostlin nemá oscilátor specifickou lokalizaci. Při řízení autonomních pohybů listů je zdrojem rytmicity pulvinus („hybný polštářek“) řapíku listu, při udržování rytmu v metabolizmu CO2 u Bryophyllum je autonomní epidermis (pokožka), ne však již mezofyl (vnitřní pletivo) listu. V souladu s hromadícími se důkazy funkce c-AMP (cyklického adenosinmonofosfátu) a inositoltrisfosfátu jako druhých poslů (látek přenášejících informaci z membrány do nitra buňky) se i u rostlin ukazuje, že tyto látky se mohou významně podílet v regulaci rytmicity. Vzhledem k tomu, že inositolový signální systém je úzce propojen s regulací hladiny vápníku v buňce, nepřekvapí nás, že i jeho hladina může ovlivňovat rytmicitu. Ostatně jak se ukázalo při studiu pohybu listů, i změny toků dalších iontů (protonů, draslíku, chloridových) mohou mít pro rytmicitu význam. Potíž je v tom, že i na úrovni druhých poslů a změn transkripce jsme stále blíže ručičkám než hodinovému stroji.
 Z houby Neurospora crassa byl nedávno izolován gen frq (frequency), jehož produkty jsou nutné k udržení rytmu konidiace a buněčného dělení. Transkripcí genu frq však vznikají dva produkty a dosud nebyl objasněn jejich vzájemný vztah. Gen frq vykazuje poměrně nízkou homologii s podobným genem z klasického objektu studia rytmicity – banánové mušky Drosophila melanogaster. Přestože to bylo vlastně trochu zklamáním, jde jistě o významný výchozí bod pro další výzkum. Hybridizačními testy byly sekvence podobné genu frq z neurospory nalezeny i v genomu huseníčku Arabidopsis thaliana, sóje, kvasinek, ale i u některých savců. Nakonec se tedy zdá, že přece jenom bude mít řízení rytmicity vysokou homologii u všech organizmů.
Z houby Neurospora crassa byl nedávno izolován gen frq (frequency), jehož produkty jsou nutné k udržení rytmu konidiace a buněčného dělení. Transkripcí genu frq však vznikají dva produkty a dosud nebyl objasněn jejich vzájemný vztah. Gen frq vykazuje poměrně nízkou homologii s podobným genem z klasického objektu studia rytmicity – banánové mušky Drosophila melanogaster. Přestože to bylo vlastně trochu zklamáním, jde jistě o významný výchozí bod pro další výzkum. Hybridizačními testy byly sekvence podobné genu frq z neurospory nalezeny i v genomu huseníčku Arabidopsis thaliana, sóje, kvasinek, ale i u některých savců. Nakonec se tedy zdá, že přece jenom bude mít řízení rytmicity vysokou homologii u všech organizmů.
Na příkladu fotoperiodicky navozeného kvetení se pokusíme s využitím předcházejících informací demonstrovat účast endogenní rytmicity na procesu kontrolovaném fytochromem.
Příjem fotoperiodického signálu
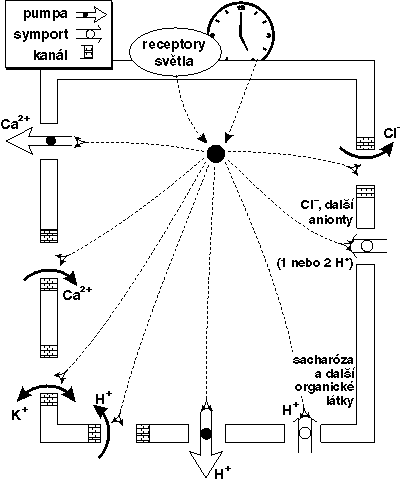
Na rozdíl od živočichů neznáme u rostlin ani specializované čidlo příjmu světelného signálu, ani analogii nervové soustavy, která by signál zpracovala. Absorpční maxima akčních spekter některých procesů kontrolovaných světlem, mezi něž patří i fotoperiodické projevy, jsou omezena pouze na určité vlnové délky. Nejčastěji jde o oblast červeného 660 nm (red, R) a dlouhovlnného červeného záření 730 nm (far red, FR). Další aktivní oblastí je světlo modré (akční spektra mívají maxima při 370, 450 a 480 nm) a také UV-záření zvané UVb v oblasti 280–320 nm. Světlo červené oblasti rostlina registruje prostřednictvím fotoreceptoru fytochromu. Jde o fotoreverzibilní chromoprotein s relativní molekulovou hmotností 124 kDa a prostetickou (funkční) skupinou tetrapyrolové struktury. Ta při absorbci světla s vlnovou délkou 660 nm mění svoji konfiguraci a vrací se k původní formě působením světla s vlnovou délkou 730 nm (obrázek). Zároveň se mění i struktura celého bílkovinného komplexu. Biologicky aktivní je forma vznikající osvětlením R a je označována Pfr je nestálá a zčásti přechází zpět na formu Pr, zčásti je degradována (obrázek). Forma a tedy i účinek fytochromu závisí na typu posledního osvětlení. K biologicky aktivní fotokonverzi (přechod Pr na Pfr) a ustavení rovnováhy Pr/Ppr postačuje světlo velmi nízké intenzity (ca 1 W.m-2).Takové světlo není fotosynteticky aktivní a lze Lak rozlišit fotoperiodické a fotomorfogenní vlivy světla od vlivů zprostředkovaných fotosyntézou. Funkci fytochromu jako příjemce světla odpovídá i jeho univerzální rozšíření v říši rostlin. Nachází se ve všech orgánech včetně kořenů, a to i u rostlin etiolovaných (bez chlorofylu). Vyskytuje se i u zelených řas, mechů a kapraďorostů a všech druhů rostlin nahosemenných a krytosemenných. Mechanizmus přenosu světelného signálu není dosud plně objasněn. Změna struktury fytochromu, jehož část a příslušný receptor jsou vázány v plazmatické membráně, má za následek změnu toku iontů, zejména vápníku, který představuje u rostlin část signálního systému. Ionty vápníku po vazbě na specifickou bílkovinu kalmodulin jsou schopny aktivovat některé enzymy, zejména proteinkinázy. Mění se membránový potenciál i toky dalších iontů, mění se i fosforylace proteinů (obrázek). Dále fytochrom reguluje expresi genomu a pod jeho kontrolou je syntéza více než 60 enzymů, ať již jde o ovlivnění na úrovni transkiripční, či translační. Jako příklad transkipční kontroly můžeme uvést syntézu mRNK (informační RNK) pro malou subjednotku hlavního fotosyntetického enzymu ribulozobisfosfátkarboxylázy-oxygenázy (Rubisco). Citlivost ke světlu lze připisovat úsekům v tzv. regulačních oblastech genů, tj. ve směru 5’ od startu transkripce, které jsou homologní u většiny genů citlivých ke světlu.
Světlo červené oblasti rostlina registruje prostřednictvím fotoreceptoru fytochromu. Jde o fotoreverzibilní chromoprotein s relativní molekulovou hmotností 124 kDa a prostetickou (funkční) skupinou tetrapyrolové struktury. Ta při absorbci světla s vlnovou délkou 660 nm mění svoji konfiguraci a vrací se k původní formě působením světla s vlnovou délkou 730 nm (obrázek). Zároveň se mění i struktura celého bílkovinného komplexu. Biologicky aktivní je forma vznikající osvětlením R a je označována Pfr je nestálá a zčásti přechází zpět na formu Pr, zčásti je degradována (obrázek). Forma a tedy i účinek fytochromu závisí na typu posledního osvětlení. K biologicky aktivní fotokonverzi (přechod Pr na Pfr) a ustavení rovnováhy Pr/Ppr postačuje světlo velmi nízké intenzity (ca 1 W.m-2).Takové světlo není fotosynteticky aktivní a lze Lak rozlišit fotoperiodické a fotomorfogenní vlivy světla od vlivů zprostředkovaných fotosyntézou. Funkci fytochromu jako příjemce světla odpovídá i jeho univerzální rozšíření v říši rostlin. Nachází se ve všech orgánech včetně kořenů, a to i u rostlin etiolovaných (bez chlorofylu). Vyskytuje se i u zelených řas, mechů a kapraďorostů a všech druhů rostlin nahosemenných a krytosemenných. Mechanizmus přenosu světelného signálu není dosud plně objasněn. Změna struktury fytochromu, jehož část a příslušný receptor jsou vázány v plazmatické membráně, má za následek změnu toku iontů, zejména vápníku, který představuje u rostlin část signálního systému. Ionty vápníku po vazbě na specifickou bílkovinu kalmodulin jsou schopny aktivovat některé enzymy, zejména proteinkinázy. Mění se membránový potenciál i toky dalších iontů, mění se i fosforylace proteinů (obrázek). Dále fytochrom reguluje expresi genomu a pod jeho kontrolou je syntéza více než 60 enzymů, ať již jde o ovlivnění na úrovni transkiripční, či translační. Jako příklad transkipční kontroly můžeme uvést syntézu mRNK (informační RNK) pro malou subjednotku hlavního fotosyntetického enzymu ribulozobisfosfátkarboxylázy-oxygenázy (Rubisco). Citlivost ke světlu lze připisovat úsekům v tzv. regulačních oblastech genů, tj. ve směru 5’ od startu transkripce, které jsou homologní u většiny genů citlivých ke světlu.
 Fytochrom se s největší pravděpodobností účastní i měření času, a tedy v regulaci všech periodických, rytmických projevů rostlin. Ve světelné fázi, jak jsme již uváděli, se tvoří aktivní forma Pfr, jejíž hladina pak ve tmě klesá. Rychlost této konverze je prakticky nezávislá na teplotě. Tak by hladina Pfr, přesněji řečeno poměrné zastoupení Pfr v celkovém „poolu“ fytochromu, mohla sloužit k měření času, k určení délky dne a noci. Tato představa je patrně zjednodušená, byť byla kdysi základem všech výkladů o úloze fytochromu v registraci fotoperiodických podmínek a bylo navrženo mnoho podstatně složitějších modelů. Všechny ale se změnami hladin aktivní formy fytochromu operují. Na významnou roli fytochromu ukazují i efekty červeného světla (R), které fáze fotoperiodicky regulovaných rytmů posouvají, nebo je ruší, jak se o tom ještě zmíníme při výkladu fotoperiodicky řízeného kvetení.
Fytochrom se s největší pravděpodobností účastní i měření času, a tedy v regulaci všech periodických, rytmických projevů rostlin. Ve světelné fázi, jak jsme již uváděli, se tvoří aktivní forma Pfr, jejíž hladina pak ve tmě klesá. Rychlost této konverze je prakticky nezávislá na teplotě. Tak by hladina Pfr, přesněji řečeno poměrné zastoupení Pfr v celkovém „poolu“ fytochromu, mohla sloužit k měření času, k určení délky dne a noci. Tato představa je patrně zjednodušená, byť byla kdysi základem všech výkladů o úloze fytochromu v registraci fotoperiodických podmínek a bylo navrženo mnoho podstatně složitějších modelů. Všechny ale se změnami hladin aktivní formy fytochromu operují. Na významnou roli fytochromu ukazují i efekty červeného světla (R), které fáze fotoperiodicky regulovaných rytmů posouvají, nebo je ruší, jak se o tom ještě zmíníme při výkladu fotoperiodicky řízeného kvetení.
 Význam fytochromu pro růst a vývoj rostlin názorně ukazují mutanty s nedostatkem fytochromu. Tyto mutanty mají často i na světle vzhled rostlin etiolovaných (vytáhlé, slabé rostliny téměř bez chlorofylu). Jiné mutanty se naopak i ve tmě vyvíjejí podobně jako na světle. Fytochrom můžeme tedy považovat za součást regulace fotoperiodicky řízených vývojových jevů, kam patří květní iniciace u bylin a výjimečně i u dřevin (např. u kávovníku), klíčení semen, tvorba hlíz (tuberizace), navození klidového období (dormance) pupenů víceletých bylin i dřevin, tvorba zimních pupenů dřevin, opad listů. Kromě toho se fytochrom podílí na řízení růstových procesů, které nesouvisí s měřením času, např. stimulace dlouživého růstu při zastínění. Tyto tzv. fotomorfogenetické efekty, jež s vývojem nesouvisí přímo, zde probírat nebudeme a v další kapitole se pokusíme demonstrovat interakci fytochromu s rytmy na příkladu fotoperiodicky regulovaného kvetení.
Význam fytochromu pro růst a vývoj rostlin názorně ukazují mutanty s nedostatkem fytochromu. Tyto mutanty mají často i na světle vzhled rostlin etiolovaných (vytáhlé, slabé rostliny téměř bez chlorofylu). Jiné mutanty se naopak i ve tmě vyvíjejí podobně jako na světle. Fytochrom můžeme tedy považovat za součást regulace fotoperiodicky řízených vývojových jevů, kam patří květní iniciace u bylin a výjimečně i u dřevin (např. u kávovníku), klíčení semen, tvorba hlíz (tuberizace), navození klidového období (dormance) pupenů víceletých bylin i dřevin, tvorba zimních pupenů dřevin, opad listů. Kromě toho se fytochrom podílí na řízení růstových procesů, které nesouvisí s měřením času, např. stimulace dlouživého růstu při zastínění. Tyto tzv. fotomorfogenetické efekty, jež s vývojem nesouvisí přímo, zde probírat nebudeme a v další kapitole se pokusíme demonstrovat interakci fytochromu s rytmy na příkladu fotoperiodicky regulovaného kvetení.
Cirkadiánní rytmus u fotoperiodicky vyvolaného kvetení
Budeme se zabývat kvetením u krátkodenního druhu merlíku červeného (Chenopodium rubrum), který je modelovou rostlinou pro studium fotoperiodické indukce a jejž jsme sami ve své práci používali (obrázek). Její přednost jako „laboratorní“ rostliny je jak ve vysoké fotoperiodické citlivosti – postačí osm hodin temnoty, aby následně vykvetl celý soubor, tak v okolnosti, že kvést mohou již malé klíční rostliny – stovky jedinců na jediné Petriho misce. Kritická délka temné periody pro kvetení i závislost fotoperiodické citlivosti na stáří jsou znázorněny na obrázku. U tohoto druhu byl zjištěn cirkadiánní endogenní rytmus kvetení, který je vyvolán přenesením rostlin ze světla do tmy. První vrchol pozorujeme po 13 hodinách a délka periody, je 30 až 31 hodin (obrázek). Vyznívání rytmu v druhém a třetím cyklu, projevující se snížením amplitudy (% kvetoucích rostlin), je spojeno s dlouhodobým setrváním rostlin v temnotě a nedostatkem energeticky bohatých substrátů, např. sacharózy.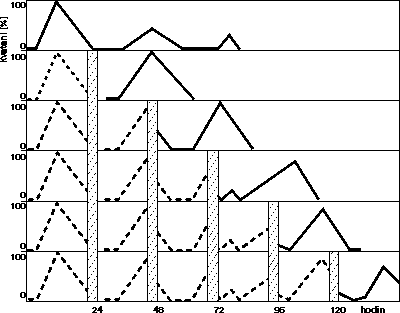 O tom, že květní indukce je řízena fytochromem, svědčí možnost anulovat působení indukční temné periody krátkodobým (5 – 20’) působením červeného světla (R). Tento efekt lze „vymazat“ účinkem dlouhovlnného červeného záření (FR), které upravuje předchozím zásahem zvýšenou hladinu Pfr na původní. Na střídání fotofilní a skotofilní fáze rytmu ukazuje změna účinnosti červeného světla, která je zachycena na obrázku. Z hlediska účasti endogenního rytmu při měření času v přirozených podmínkách mají zvláštní význam opakované vyvolání rytmu při každém přechodu světlo/tma a fázový posun spontánní podoby rytmu v závislosti na délce temné periody. Na modelovém příkladu, který uvádíme, bylo použito cyklů šesti hodin světla a osmnácti hodin temnoty. K fázovému posunu, tj. k urychlení nebo zpomalení rytmu vlivem fotoperiody, dochází v závislosti na fázi rytmu fotoperiodou zasažené. Na počátku temnoty působí fotoperioda zpoždění rytmu, v pozdější fázi urychlení. Jak vyplývá z (obrázku, který ukazuje několik cyklů s postupným refázováním rytmu, teprve po pěti cyklech se endogenní rytmus synchronizuje s fotoperiodickými podmínkami, jimž je rostlina vystavena. Tím bylo dosaženo koincidence mezi průběhem spontánní fáze rytmu a fotoperiodou prostředí a „hodinový stroj“ rostliny je připraven měřit nebeský čas.
O tom, že květní indukce je řízena fytochromem, svědčí možnost anulovat působení indukční temné periody krátkodobým (5 – 20’) působením červeného světla (R). Tento efekt lze „vymazat“ účinkem dlouhovlnného červeného záření (FR), které upravuje předchozím zásahem zvýšenou hladinu Pfr na původní. Na střídání fotofilní a skotofilní fáze rytmu ukazuje změna účinnosti červeného světla, která je zachycena na obrázku. Z hlediska účasti endogenního rytmu při měření času v přirozených podmínkách mají zvláštní význam opakované vyvolání rytmu při každém přechodu světlo/tma a fázový posun spontánní podoby rytmu v závislosti na délce temné periody. Na modelovém příkladu, který uvádíme, bylo použito cyklů šesti hodin světla a osmnácti hodin temnoty. K fázovému posunu, tj. k urychlení nebo zpomalení rytmu vlivem fotoperiody, dochází v závislosti na fázi rytmu fotoperiodou zasažené. Na počátku temnoty působí fotoperioda zpoždění rytmu, v pozdější fázi urychlení. Jak vyplývá z (obrázku, který ukazuje několik cyklů s postupným refázováním rytmu, teprve po pěti cyklech se endogenní rytmus synchronizuje s fotoperiodickými podmínkami, jimž je rostlina vystavena. Tím bylo dosaženo koincidence mezi průběhem spontánní fáze rytmu a fotoperiodou prostředí a „hodinový stroj“ rostliny je připraven měřit nebeský čas.
 Znalost těchto formálních zákonitostí průběhu rytmických projevů ještě nijak nepřispívá k poznání procesu, jehož časování je rytmicitou řízeno. V našem případě víme, že sledovaný proces je lokalizován v dělohách či listech a že je spojen s vytvářením chemického signálu kvetení – florigenního stimulu(-ů). Ten je transportován vodivými svazky rychlostí cca 15–20 mm.h-1 do vrcholu, kde vyvolává nový program morfogenní aktivity – namísto listů se tvoří květy. Ale studium mechanizmů květní indukce je jiná kapitola; nás bude nyní zajímat, jaký je ekologický, adaptační význam rytmicity fotoperiodicky vyvolaného kvetení.
Znalost těchto formálních zákonitostí průběhu rytmických projevů ještě nijak nepřispívá k poznání procesu, jehož časování je rytmicitou řízeno. V našem případě víme, že sledovaný proces je lokalizován v dělohách či listech a že je spojen s vytvářením chemického signálu kvetení – florigenního stimulu(-ů). Ten je transportován vodivými svazky rychlostí cca 15–20 mm.h-1 do vrcholu, kde vyvolává nový program morfogenní aktivity – namísto listů se tvoří květy. Ale studium mechanizmů květní indukce je jiná kapitola; nás bude nyní zajímat, jaký je ekologický, adaptační význam rytmicity fotoperiodicky vyvolaného kvetení.
Ekologický význam rytmicity fotoperiodicky řízeného kvetení
Obecný adaptační význam endogenní rytmicity lze spojovat se synchronizováním vnitřních procesů rostliny s cyklickými projevy prostředí. Endogenní rytmicita pří tom vytváří pozadí pro působení signálů prostředí. Ať již jde o optimalizaci denního průběhu funkcí (adaptaci na dlouhodobé změny přírůstku a úbytku délky dne), nebo sezonního průběhu vývoje. Druhý případ zahrnuje i iniciaci kvetení. Iniciace kvetení, řízená fotoperiodickou reakcí a endogenní rytmicitou, je v temperátní zóně časována tak, aby byla vyloučena rizika nepříznivých podmínek zimy (fotoperiodické podmínky „zabraňují“ kvetení) a naopak bylo „využito“ pro tvorbu semen a plodů období s maximálním příkonem sluneční energie. Významným adaptačním rysem tedy je schopnost predikce průběhu sezonního počasí. Proto v naší zeměpisné šířce převažují dlouhodenní rostliny a proto je květen v květnu, před nástupem dlouhých dnů a zvýšených teplot časného léta.Omluvme se za poněkud teleologický tón výkladu a skončeme konstatováním, že fytogeografický a fytocenologlcký význam fotoperiodické iniciace kvetení se odráží i ve skutečnosti, že se dlouhodenní fotoperiodická indukce kvetení předpokládá u zhruba 80 našich planě rostoucích bylinných druhů. Na druhé straně je fotoperiodicky regulován nástup reproduktivní fáze i u našich klíčových hospodářských plodin: pšenice, ječmene a cukrové řepy i brambor. Argument nejen pro estéty, ale i pro jedlíky, aby oblast zůstala badatelsky aktivní.
Citát
Carl Friedrich von Weizsäcker při příležitosti udělení ceny Karla IV.
Moderní věda umožnila moderní techniku. Technika vytváří rychlý přírustek populace a tím nové celosvětové sociální konflikty. Technika mění naše fyzické prostředí: jak bude vypadat klima za několik desetiletí? Lidstvo bude moci žít s technikou jedině tehdy, bude-li zavčas důsledky techniky promýšlet a odpovídajícím způsobem jednat. Banální příklad: jsou-li možno auta, je nutný řád silničního provozu. Ekonomický příklad: teorie tržního hospodářství od Adama Smíthe se týkala „wealth of nations“, blahobytu uvnitř obroda. Dnes je trh světovým trhem: Potřeboval by právní a sociální regulaci, kterou ještě nemá.
Troufám si tvrdit: neexistuje žádný technický a sociální problém, který by nebyl řešitelný pomocí společně používaného rozumu. Myslet na toto používání rozumu a učit ho, to je odpovědnost vědy. Každý rozumný politik v demokracii udělá, co je rozumná, bude-li vědět, že příští volby neprohraje, jak se ještě často stává, protože učinil, co bylo rozumné. Zde musí pomáhat věda ve vytváření veřejného mínění. Historická odpověď: člověk vzešel z evoluce. Evoluce trvala mnoho milionů let. Pět tisíciletí je krátká doba. Člověk je učenlivá bytost. Kolik se toho za těchto několik tisíciletí naučil! Existovala města. Existovala knížectví. Existovaly velké státy. Uvnitř měst bylo již záhy zakázáno, aby jeden občan zabíjel druhého. Ve velkých státech byly zákony prosazovány prostřednictvím vládnoucí moci. Že se občané různých států nemají navzájem zabíjet, že je třeba překonat volku jako instituci, počíná tušit naše století.
Ve skutečnosti je toto poznání staré již celo tisíciletí. Velká náboženství učila hlubší, reálnější etice: židovství, křesťanství, hinduizmus, buddhizmus. Římský císař Konstantin udělal z křesťanství státní náboženství, zřejmě proto, že křesťané byli ochotni za svou etiku zemřít, byli jedinou silnou, spolehlivou minoritou ve zmatené říši. Ovšem když vládli, propadli mocenská víře jako jejich římští předchůdci. Bylo zapotřebí vnitřních křesťanských reformních hnutí, jako např. reforem františkánů. Byli i husité takovou korekturou? Náboženství je včasný hluboký náhled do lidských motivů a je vůlí propůjčit dobrým motivům účinek. Věda je osvěta. Učí pochopení, umožňuje techniku, s níž spojuje nepostradatelnost rozumu. Troufám si říci: náboženství se dnes přiblíží svému cíli pouze tehdy, vezme-li tuto osvětu naprosto vážně. Osvěta se přiblíží svému cíli pouze tehdy, vezme-li naprosto vážně náboženství. Je dost možné, že tato cesta povede přece jen ještě bezpříkladnými velkými katastrofami. Ale je to cesta.
ZÁKLADNÍ POJMY PRO VÝKLAD RYTMICITY
Amplituda: poloviční vzdálenost od minimální k maximální hodnotě rytmicky se měnící veličiny.
Cyklus: jestliže se daný proces vrací do výchozího stavu, mluvíme o cyklu.
Časovač: frekvence endogenního rytmu se dá uvést do souladu s nějakou vnější oscilací, např. střídáním dne a noci. Fáze rytmuje pak posouvána, zrychlována či zpomalována signálem od takové oscilace odvozeným, jemuž říkáme časovač (dle Bünninga Zeitgeber) či synchronizátor. Může jím být např. přechod ze světla do tmy a naopak, tj. v přírodě stmíváni a sultáni.
Fáze: určitý (libovolný) časový bod od počátku cyklu – po uplynuti jedné periody se rytmus vždy nachází ve stejné fázi.
Frekvence: počet cyklů v daném čase. Perioda: doba trváni jednoho cyklu. Spontánní rytmus: rytmus, který nezávisí na vnějších podmínkách a v konstantních podmínkách vykazuje svou přirozenou periodu.
Zavedeni či vzbuzení rytmu: rytmické chování sledovaného jevu je zaváděno nějakým signálem, u fotoperiodických dějů často např. přechodem ze světla do tmy.
O autorech
Jan Krekule
Ivana Macháčková
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
