










O čem vypovídají chloroplasty sítin
Už od dob Charlese Darwina je rekonstrukce evolučních dějin všech organizmů na Zemi a její vyjádření v podobě fylogenetického stromu snem mnoha biologů.
Čeleď sítinovité v současné době zahrnuje 442 druhů členěných do sedmi rodů. Dva z nich, sítina (Juncus) a bika (Luzula), se vyskytují po celém světě, ostatních pět (Rostkovia, Oxychloë, Patosia, Distichia a Marsippospermum) 1) najdeme převážně v Jižní Americe (obrázek 1). Na první pohled se sítinovité rostliny podobají travám, ale liší se od nich řadou významných znaků, jako je např. nepřítomnost stébel s kolénky nebo stavba květu. Také od blízce příbuzných šáchorovitých (Cyperaceae) rozeznáte sítinovité na první pohled podle vícesemenných tobolek, semeníku srostlého ze tří plodolistů a většinou oblé lodyhy.
Zabývali jsme se studiem fylogenetických vztahů uvnitř čeledi sítinovitých za použití molekulárních dat, zejména sekvencí chloroplastové DNA (cpDNA). Chloroplastový genom je totiž poměrně malý (120–200 kb), což usnadňuje jeho analýzu (u několika převážně modelových či ekonomicky významných rostlin již je známa sekvence celého chloroplastového genomu). Další výhodou je poměrně pomalá evoluce cpDNA; důležité ovšem je, že se různé části cpDNA vyvíjely různou rychlostí, což poskytuje spoustu možností pro řešení vztahů mezi rostlinami od úrovně druhu a rodu až po čeleď. Chloroplasty jsou navíc v rostlinných buňkách přítomny v mnoha kopiích, což zvyšuje výtěžek DNA při izolaci a také pravděpodobnost, že i v herbářovém materiálu vydrží alespoň část cpDNA v použitelném stavu.
Jak se staví fylogenetický strom
 Prvním krokem ke konstrukci fylogenetického stromu je vždy získání primárních dat. My jsme vycházeli především ze sekvencí tří úseků cpDNA. Prvním z nich byl gen pro velkou podjednotku fotosyntetického enzymu ribulózobisfosfátkarboxylázy (RuBisCo) – rbcL, zbylé dva odpovídají nekódujícím oblastem (intron trnL a mezigenový „mezerník“ trnL-F). Ve všech případech jsme sledovali jednak bodové mutace, jednak vzácné strukturní přestavby (inzerce a delece), 2) které bývají považovány za doklad společného původu svých nositelů.
Prvním krokem ke konstrukci fylogenetického stromu je vždy získání primárních dat. My jsme vycházeli především ze sekvencí tří úseků cpDNA. Prvním z nich byl gen pro velkou podjednotku fotosyntetického enzymu ribulózobisfosfátkarboxylázy (RuBisCo) – rbcL, zbylé dva odpovídají nekódujícím oblastem (intron trnL a mezigenový „mezerník“ trnL-F). Ve všech případech jsme sledovali jednak bodové mutace, jednak vzácné strukturní přestavby (inzerce a delece), 2) které bývají považovány za doklad společného původu svých nositelů.
Geny, jako třeba rbcL, jsou si většinou vzájemně natolik podobné, že můžeme snadno určit příbuzné (homologní) sekvence (viz Zrzavý, Vesmír 76, 27, 1997). Vzájemným přiřazením sekvencí rbcL tedy dostaneme matici dat s větším či menším množstvím substitucí, a z počtu substitucí můžeme usoudit na míru příbuznosti zkoumaných taxonů. U nekódujících úseků se příbuzné úseky hledají hůře, a konstrukce přiřazení je obtížnější, protože v průběhu evoluce dochází k řadě přestaveb (obrázek 2).
 Na přiřazení sekvencí navazuje vlastní konstrukce fylogenetického stromu některým ze standardních algoritmů a statistické hodnocení. Výsledkem je vlastní fylogenetický strom, zahrnující i údaje o pravděpodobnosti jednotlivých větví a další statistické informace. 3)
Na přiřazení sekvencí navazuje vlastní konstrukce fylogenetického stromu některým ze standardních algoritmů a statistické hodnocení. Výsledkem je vlastní fylogenetický strom, zahrnující i údaje o pravděpodobnosti jednotlivých větví a další statistické informace. 3)
Co nám řeknou sekvenční přestavby: případ pseudogenů
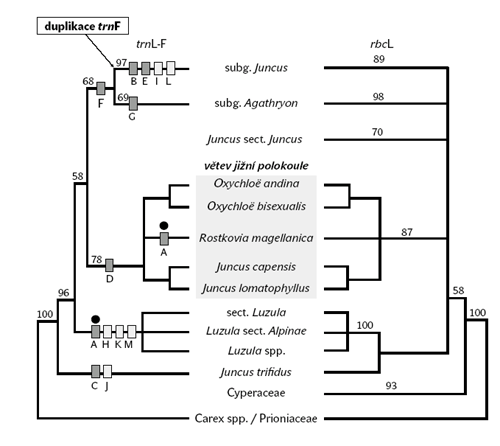
Pseudogeny jsou nefunkční úseky DNA, jejichž sekvence se podobá „normálním“ funkčním genům. Dosud známe dva mechanizmy vzniku pseudogenů – zaprvé zdvojení úseku DNA s následným rozrušením „nadbytečné“ kopie genu mutacemi, zadruhé retrotranspozici, při které se do genomu vloží fragment DNA vzniklý zpětným přepisem molekuly RNA (viz Vesmír 79, 273, 2000/5). Pseudogeny byly nalezeny u všech modelových organizmů od kvasinek přes huseníček až po člověka. Z hlediska evolučních studií jsou pseudogeny důležité i proto, že umožňují sledovat evoluci sekvencí DNA a určovat rychlost vzniku mutací bez selekčního tlaku.I u našich sítin byl nalezen pseudogen trnF, který je zřejmě typický jen pro sítiny a u bik a dalších rodů čeledi se vůbec nevyskytuje, ba dokonce je specifický jen pro podrod Juncus (viz obrázek 5), což lze pokládat za nezávislé potvrzení oprávněnosti tohoto podrodu.
Případ Oxychloë: zápletka přímo detektivní
Rod Oxychloë je malý endemický rod s 5 druhy; centrem výskytu jsou vysoké Andy Peru, Bolívie, severní Chile a Argentiny. Na základě morfologických znaků je tento rod zařazován mezi sítinovité, do blízkosti rodu Patosia, od něhož se odlišuje pouze přítomností květů se dvěma listeny (Patosia má jen jeden listen) a tvarem listové čepele (zubatý okraj oproti hladkému). Nic by tedy nenasvědčovalo, že by tento rod mohl být nějak problematický. Překvapení přineslo až sekvencování chloroplastového genu rbcL Oxychloë andina (obrázek 3) dvěma nezávislými skupinami v letech 1993 a 1995. Porovnání s dalšími zástupci čeledi sítinovitých a šáchorovitých totiž ukázalo, že druh Oxychloë patří mezi šáchorovité! Autoři sekvencí usoudili, že „sítinovité“ morfologické znaky jsou zřejmě původní, a dál se záhadou nezabývali.
Překvapení přineslo až sekvencování chloroplastového genu rbcL Oxychloë andina (obrázek 3) dvěma nezávislými skupinami v letech 1993 a 1995. Porovnání s dalšími zástupci čeledi sítinovitých a šáchorovitých totiž ukázalo, že druh Oxychloë patří mezi šáchorovité! Autoři sekvencí usoudili, že „sítinovité“ morfologické znaky jsou zřejmě původní, a dál se záhadou nezabývali.
I naše analýzy vycházející z publikovaných sekvencí rbcL potvrdily zařazení Oxychloë mezi šáchorovité. Nechtěli jsme však konstrukci fylogenetického stromu čeledi sítinovitých založit jen na jednom genu, a proto jsme vybrali další, totiž dvě nekódující oblasti chloroplastu, intron trnL a mezerník trnL-F. Tentokrát však patřičné sekvence z Oxychloë nebyly v databázi zastoupeny. Následovalo pátrání po živém či alespoň herbářovém materiálu, což se ukázalo jako velký problém, neboť tento druh poměrně málo kvete, je tedy velmi často sbírán jako sterilní a těžko se určuje. Nakonec jsme získali a analyzovali zástupce čtyř z pěti druhů (O. andina, O. bisexualis, O. castellanosii a O. haumaniana) a s údivem jsme zjistili, že všechny čtyři druhy vycházejí zařazeny mezi sítinovité, a dokonce „správně“ do větve s dalšími druhy jižní polokoule. Pro další doplnění a upřesnění vztahů uvnitř skupiny jsme analyzovali další chloroplastový gen i vybrané jaderné sekvence. 4) Všechna data prokázala, že rod Oxychloë je skutečně zástupce sítinovitých.
Jak je možné, že se naše data tak zásadně liší od sekvencí v databázi? Po prohlédnutí jedné z herbářových položek, z níž byl odebrán materiál na analýzu DNA, jsme zjistli, že jde o směsnou položku, kde je skutečně zastoupen druh Oxychloë andina (dokonce v plodném stavu!), ale i jiná, neplodící tráva, z níž byl prokazatelně také odebírán materiál. Druhá sporná herbářová položka skutečně patřila druhu O. andina. Izolovali jsme DNA z této rostliny a získali sekvenci, která již není v analýze řazena mezi šáchorovité, nýbrž mezi sítinovité. U sekvence z databáze došlo tedy zcela jistě v průběhu analýz ke kontaminaci DNA z jiné rostliny. Tak tedy konec záhady, Oxychloë bezesporu opět patří mezi sítinovité.
Čím je zvláštní sítina trojklaná?
 Chloroplastová data ukazují sítinu trojklanou jako původní taxon pro rod bika (rbcL data), nebo dokonce pro celý zbytek čeledi sítinovitých, včetně malých jihoamerických rodů (trnL-F). Toto zjištění není zas až tak překvapivé, neboť již G. Rouy na počátku 20. století (r. 1912) stanovil samostatnou sekci Trifidii na základě morfologické výjimečnosti, zahrnující sítinu trojklanou a blízce příbuznou sítinu jednokvětou (Juncus monanthos). Oba tyto druhy mají skutečně unikátní vzhled (obrázek 4). Terminální redukované květenství je tvořeno 1–3 květy, které jsou převýšeny listenem.
Chloroplastová data ukazují sítinu trojklanou jako původní taxon pro rod bika (rbcL data), nebo dokonce pro celý zbytek čeledi sítinovitých, včetně malých jihoamerických rodů (trnL-F). Toto zjištění není zas až tak překvapivé, neboť již G. Rouy na počátku 20. století (r. 1912) stanovil samostatnou sekci Trifidii na základě morfologické výjimečnosti, zahrnující sítinu trojklanou a blízce příbuznou sítinu jednokvětou (Juncus monanthos). Oba tyto druhy mají skutečně unikátní vzhled (obrázek 4). Terminální redukované květenství je tvořeno 1–3 květy, které jsou převýšeny listenem.
Co se týče molekulárních odlišností, sítina trojklaná je charakterizována dvěmi strukturními přestavbami, delecí v intronu trnL a inzercí v mezerníku trnL-trnF (obrázek 2, obrázek 5). Obě přestavby jsou zcela jedinečné mezi druhy rodu sítina i bika. Oprávněnost samostatnosti sekce Trifidii byla tedy potvrzena i na základě molekulárních dat.
Strom genů, nebo druhů?
Z případu sítiny trojklané by se zdálo, že nám nyní již nic nebrání obnovit statut sekce Trifidii či udělat jiné taxonomicky závažné změny. Fylogenetický strom bývá však často konstruován z jednoho nebo několika málo úseků DNA pro každý druh, a nemusí nutně souhlasit s fylogenezí druhu. Časové větvení mezi dvěma druhy u stromu druhu totiž ukazuje dobu reprodukční izolace daných druhů, oproti tomu genový strom ukazuje dobu „větvení“ daného genu či genové rodiny. Nelze tedy měnit taxonomii dané skupiny na základě dat z jednoho jediného úseku genomu, i když se to v prvotním nadšení nad aplikací molekulárněbiologických metod do oblasti systematiky mnohdy dělo. Pro skupinu blízce příbuzných druhů nebo populací však často genový strom nesouhlasí s druhovým, jak ukázaly například studie cichlid z afrického jezera Victoria nebo mitochondriální DNA různých lidských populací.Do fylogenetické analýzy je tedy nutné začlenit úseky ze všech třech genomů (jádra, mitochondrie, chloroplastu) a molekulární data porovnávat i s anatomicko-morfologickým rozborem.
Co víme nového o evoluci sítinovitých
 Jedním z nejvýznamnějších výsledků naší práce bylo zjištění, že všechny biky, které dnes vidíme kolem sebe třeba na louce (bika ladní – Luzula campestris) nebo v lese (bika lesní – Luzula sylvatica), jsou potomky jednoho předka, „pramáti biky“, tedy výsledkem jedné evoluční linie. Toto zjištění je velmi důležité, neboť struktura fylogenetického stromu ukazuje, že rod se rozrůznil až po oddělení kontinentů; můžeme tedy vysledovat skupiny druhů vymezené i místem výskytu (např. jihoamerické biky Luzula peruviana a L. racemosa).
Jedním z nejvýznamnějších výsledků naší práce bylo zjištění, že všechny biky, které dnes vidíme kolem sebe třeba na louce (bika ladní – Luzula campestris) nebo v lese (bika lesní – Luzula sylvatica), jsou potomky jednoho předka, „pramáti biky“, tedy výsledkem jedné evoluční linie. Toto zjištění je velmi důležité, neboť struktura fylogenetického stromu ukazuje, že rod se rozrůznil až po oddělení kontinentů; můžeme tedy vysledovat skupiny druhů vymezené i místem výskytu (např. jihoamerické biky Luzula peruviana a L. racemosa).
U sítin se naopak zdá, že prapředků bylo více, podobně jako třeba u nahosemenných rostlin (jen na základě chloroplastového genomu ale nemůžeme vytvářet obecné pravdy!). Rod Juncus tedy možná zahrnuje rostliny, které spojujeme jen z nedostatku znaků, jež by stačily na vymezení dalších skupin. Tradičně bývá tento rod rozdělován na dva podrody, Agathryon (= Poiophylli) a Juncus, které se liší přítomností či nepřítomností páru listenů v květenství a v kladogramu tvoří samostatné větve (obrázek 5). Ne všichni zástupci se ale do těchto podrodů začleňují – mimo zůstává třeba již zmíněná sítina trojklaná nebo skupina druhů z jižní polokoule. Podobné případy jsou ostatně známy u více rostlinných čeledí.
Studium čeledi sítinovitých intenzivně pokračuje i nadále, s cílem získat důkladnou fylogenezi čeledi a osvětlit evoluční zákonitosti této skupiny rostlin, z nichž zde byl nastíněn jen zlomek.
Článek vznikl na základě spolupráce s RNDr. Janem Kirschnerem, CSc., a RNDr. Čestmírem Vlčkem, Ph.D., v rámci grantového projektu GA UK (grant č. 109/2002/B-BIO/PrF) a GA ČR (grant č. 206/02/0355).
Poznámky
Ke stažení
 článek ve formátu pdf [983,07 kB]
článek ve formátu pdf [983,07 kB]
O autorovi
Lenka Záveská Drábková
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
