









Alternatívne stratégie replikácie telomér II.
V prvej časti sme naznačili, aký význam majú teloméry pri replikácii lineárnych molekúl DNA a aké následky majú zmeny v ich stabilite na procesy bunkového starnutia a nádorovej transformácie. Našou ambíciou bolo poukázať na to, že udržiavanie telomér zabezpečuje viacero navzájom zastupiteľných mechanizmov. Táto skutočnosť značne komplikuje pôvodne jednoduché predstavy o využití telomér ako terča v protinádorovej terapii. Druhá časť príspevku sa zaoberá ďalšími príkladmi riešenia replikácie a stabilizácie koncov lineárnych molekúl DNA a ich klinickými a evolučnými implikáciami.
Lekcie z alternatívnych riešení problému replikácie telomér
Je možné, že ani potlačenie všetkých mechanizmov replikácie telomér nemusí byť úspešným protinádorovým prostriedkom. Nádorová bunka pod selekčným tlakom je v podobnej situácii, ako lineárne DNA–genofory rôznych druhov organizmov, ktoré „neprišli“ na učebnicové stratégie replikácie telomér. Ich existencia je dôkazom, že riešení telomérového problému je oveľa viac, ako sme spomenuli v prvej časti.Tom Cech a nezávisle od neho aj Fuyuki Ishikawa z Tokijskéj techniky sa pokúsili napodobniť evolúciu alternatívnych mechanizmov replikácie telomér v laboratórnych podmienkach. Ich študenti geneticky vypli u kvasinky Schizosaccharomyces pombe telomerázovú i rekombinačnú dráhu replikácie telomér (u kvasiniek sa pravdepodobne netvoria t-slučky). Tak vlastne navodili situáciu, v ktorej by sa vyskytla nádorová bunka pod kombinovaným vplyvom (zatiaľ hypotetických) inhibítorov. Genetický blok nebol smrteľný a na Petriho miske sa objavilo zopár kolónií. Analýza niekoľkých z nich ukázala, že prežívajúce kvasinkové bunky sa vysporiadali s problémom replikácie koncov chromozómov tým najtriviálnejším spôsobom: aby ho nemuseli riešiť, konce každého chromozómu spojili, a tým vytvorili kružnicové molekuly.
Alternatívne modely replikácie telomér však nie sú viazané iba na laboratórne mutanty. Príroda nám ich ponúka ako prirodzené produkty darwinovskej evolúcie. Zaujímavosťou je, že jedným z nich je práve ovocná muška Drosophila melanogaster, u ktorej H. J. Müller definoval pojem teloméra. Drozofila nevyužíva ani jednu zo spomínaných stratégií replikácie telomér. Jej teloméram chýbajú akékoľvek náznaky krátkych repetícií. Harald Biessmann z Kalifornskej univerzity v Irvine a Jim Mason z National Institutes of Environmental and Health Sciences v Research Triangle Park v Severnej Karolíne zistili, že teloméry drozofily pozostávajú z mozaiky pomerne dlhých transpozónov (mobilných genetických elementov) dvoch typov (Het-A a TART). Skracovanie telomér je kompenzované sporadickým pridávaním transpozónov na konce chromozómov.
Zaujímavé je aj to, že D. melanogaster je jediným druhom v rode Drosophila, ktorý využíva „transpozónovú taktiku“ udržiavania telomér. Biessmann zistil, že ostatné druhy drozofíl majú komplexné dlhé tandemové repetície, ktoré sa pravdepodobne udržiavajú prostredníctvom homologickej rekombinácie. Takto sa radia k pakomárom Chironomus, moskytom Anopheles gambiae a rastlinám z rodov Allium, Aloe, Nothoscordum a Tulbaghia, ktoré majú podobnú organizáciu telomér.
Lineárne mitochondriálne genómy: ďalšie časti telomérovej mozaiky
Doteraz sme sa zaoberali trikmi, ktoré na replikáciu a stabilizáciu svojich koncov využívajú jadrové chromozómy. Problém údržby telomér sa však netýka iba jadra eukaryotickej bunky. „Trpia“ ním všetky lineárne DNA-genómy, bez ohľadu na ich pozíciu na fylogenetickom strome či umiestnenie v bunke. Vhodným modelom na štúdium alternatívnych dráh sú aj rôzne ďalšie genofory, ktoré sú v bunke prítomné vo forme lineárnych molekúl DNA (lineárne chromozómy viacerých druhov baktérií, niektoré plazmidy a vírusy). Sám J. D. Watson demonštroval problém replikácie telomér na príklade lineárneho DNA–genómu bakteriofága T7. DNA-vírusy s lineárnymi genómami boli v riešení problému replikácie ich koncov obzvlášť vynaliezavé: bakteriofág λ vytvára kružnicovú molekulu DNA spojením koncových sekvencií; DNA poxvírusov, ktoré majú na koncoch vlásenkovú štruktúru, sa replikuje prostredníctvom kružnicových medziproduktov; DNA-polymeráza adenovírusov využíva ako primer hydroxylovú skupinu aminokyseliny serínu, ktorá je súčasťou špecifického proteínu kovalentne pripojeného ku koncom DNA. Všetky spomenuté príklady predstavujú alternatívne stratégie riešenia toho istého problému.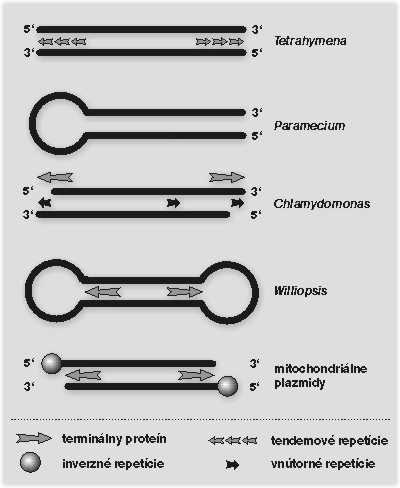 Podobná variabilita stratégií replikácie telomér bola v posledných rokoch odhalená aj u mitochondriálnych genómov. Nie, nechceme zmiasť čitateľa. Cirkularita mitochondriálnej DNA (mtDNA) je v základných učebniciach molekulárnej biológie opisovaná ako všeobecná vlastnosť mitochondriálnych genómov. Táto fyzikálna charakteristika mtDNA sa často používa ako argument v prospech endosymbiotickej teórie vzniku mitochondrií. Podľa tejto hypotézy boli predkami dnešných mitochondrií bakteriálne bunky s kružnicovou DNA. Jej podstatná časť bola v evolúcii eukaryotickej bunky eliminovaná, resp. prenesená do jadrovej DNA hostiteľa. Do väčšiny učebníc sa však z nejasných dôvodov nedostávajú zmienky o existencii lineárnych mitochondriálnych genómov. Určite to nie je pre ich zriedkavosť. Lineárne mtDNA prvokov Tetrahymena a Paramecium odhalil už na prelome 60. a 70. rokov Yoshitaka Suyama Pensylvánskej univerzity vo Filadelfii a Donald J. Cummings Coloradskej univerzity v Denveru. Začiatkom 80. rokov Micheline Wésolowská a Hiroshi Fukuhara (Paríž-Orsay) i Ladislav Kováč (v laboratóriu Piotra Slonimského v Gif-sur-Yvette) identifikovali dva druhy kvasiniek (Williopsis mrakii a Candida parapsilosis) s lineárnym mitochondriálnym genómom. Nasledujúca rozsiahla analýza vo Fukuharovom laboratóriu ukázala, že mitochondrie viac ako 20 % druhov analyzovaných kvasiniek (predovšetkým z rodov Pichia, Williopsis a Candida) majú lineárny genóm s definovanými koncovými štruktúrami – mitochondriálnymi telomérami. Zoznam organizmov s lineárnou mtDNA stále rastie a okrem prvokov a kvasiniek obsahuje aj riasy i zástupcov mnohobunkových organizmov.
Podobná variabilita stratégií replikácie telomér bola v posledných rokoch odhalená aj u mitochondriálnych genómov. Nie, nechceme zmiasť čitateľa. Cirkularita mitochondriálnej DNA (mtDNA) je v základných učebniciach molekulárnej biológie opisovaná ako všeobecná vlastnosť mitochondriálnych genómov. Táto fyzikálna charakteristika mtDNA sa často používa ako argument v prospech endosymbiotickej teórie vzniku mitochondrií. Podľa tejto hypotézy boli predkami dnešných mitochondrií bakteriálne bunky s kružnicovou DNA. Jej podstatná časť bola v evolúcii eukaryotickej bunky eliminovaná, resp. prenesená do jadrovej DNA hostiteľa. Do väčšiny učebníc sa však z nejasných dôvodov nedostávajú zmienky o existencii lineárnych mitochondriálnych genómov. Určite to nie je pre ich zriedkavosť. Lineárne mtDNA prvokov Tetrahymena a Paramecium odhalil už na prelome 60. a 70. rokov Yoshitaka Suyama Pensylvánskej univerzity vo Filadelfii a Donald J. Cummings Coloradskej univerzity v Denveru. Začiatkom 80. rokov Micheline Wésolowská a Hiroshi Fukuhara (Paríž-Orsay) i Ladislav Kováč (v laboratóriu Piotra Slonimského v Gif-sur-Yvette) identifikovali dva druhy kvasiniek (Williopsis mrakii a Candida parapsilosis) s lineárnym mitochondriálnym genómom. Nasledujúca rozsiahla analýza vo Fukuharovom laboratóriu ukázala, že mitochondrie viac ako 20 % druhov analyzovaných kvasiniek (predovšetkým z rodov Pichia, Williopsis a Candida) majú lineárny genóm s definovanými koncovými štruktúrami – mitochondriálnymi telomérami. Zoznam organizmov s lineárnou mtDNA stále rastie a okrem prvokov a kvasiniek obsahuje aj riasy i zástupcov mnohobunkových organizmov.
Existencia organizmov s lineárnou mtDNA neznamená zásadný zásah do logiky endosymbiotickej teórie. Hoci je známy nezanedbateľný počet bakteriálnych druhov s lineárnym genómom DNA (napr. niektoré druhy z rodov Borrelia a Streptomyces), a-proteobaktérie (pravdepodobní predchodcovia mitochondrií) mali kružnicovú DNA. K jej linearizácii podľa všetkého dochádzalo náhodne, ako o tom svedčí pohľad na umiestnenie organizmov s lineárnym, resp. kružnicovým mitochondriálnym genómom na fylogenetickom strome. Fukuhara dokonca odhalil dva kmene toho istého kvasinkového druhu (Williopsis suaveolens), z ktorých jeden má kružnicovú a druhý lineárnu mtDNA.
Dôsledkom náhodnej linearizácie mtDNA v rôznych fylogenetických vetvách boli odlišné spôsoby vysporiadania sa s novým problémom: replikáciou a stabilizáciou koncov – mitochondriálnych telomér. Tie sú hlavným znakom lineárnych mitochondriálnych genómov, nimi sa odlišujú od lineárnych fragmentov rôznych veľkostí prítomných vo vzorkách mtDNA kvasiniek s kružnicovým genómom. Tieto fragmenty pravdepodobne vznikajú ako produkt replikácie mtDNA mechanizmom valivej kružnice (rolling-circle; hlavnými protagonistami v tejto oblasti sú Desmond Clark-Walker z Australskej národnej univerzity a Arnold Bendich z Washingtonovy univerzity v Seattli a na stránkach Vesmíru o nej podrobnejšie písala Fatima Cvrčková (Vesmír 73, 428, 1994/8).
Vyriešenie problému replikácie koncov lineárnych mtDNA je pre príslušné druhy organizmov úlohou esenciálneho významu. Pre väčšinu by strata mitochondriálneho genómu a s ním spojená strata respiračných funkcií (mtDNA kóduje niekoľko komponentov dýchacieho reťazca) predstavovala vážne ťažkosti. Pravdaže najjednoduchším (a možno najčastejším) riešením bola opätovná cirkularizácia genómu, ako to predviedli Cechove a Ishikawove kvasinkové mutanty. Druhou alternatívou bolo „vybavenie“ koncov DNA špeciálnou telomerickou štruktúrou, ktorú môže replikovať existujúci proteínový aparát, schopný zabezpečiť kompletnú syntézu mtDNA a súčasne jej stabilizáciu i ochranu proti účinkom degradačných enzýmov. Zámerne píšeme o „už existujúcom“ proteínovom aparáte. Bunka s náhodne linearizovaným mitochondriálnym genómom nemá čas „vymýšľať“ špeciálne prostriedky replikácie telomér. V duchu jacobovského evolučného „kutilstva“ (angl. tinkering, franc. bricolage, pozri Jacob F.: Le jeu des possibles, český preklad Hra s možnostmi, Karolinum, Praha 1999) musí využiť staré prostriedky na riešenie nových problémov. K jednému konkrétnemu príkladu tohto princípu sa o chvíľu dostaneme.
Dnes poznáme viacero rôznych typov mitochondriálnych telomér, ktoré predstavujú rôzne stratégie ich replikácie (obrázek). Niektoré pripomínajú organizáciu jadrových telomér, iné teloméry adenovírusov či poxvírusov. Jeden typ mitochondriálnych telomér reprezentovaný niektorými druhmi kvasiniek (Candida parapsilosis, Candida salmanticensis a Pichia philodendra) predstavuje unikátnu organizáciu koncov lineárnych DNA-genómov. Mitochondriálne teloméry týchto kvasiniek sú tvorené dlhými (738 bázových párov Candida parapsilosis, 110 bp Candida salmanticensis, resp. 286 bp Pichia philodendra), tandemovo sa opakujúcimi sekvenciami. Vzhľadom na to, že počet týchto opakovaní nie je vo všetkých molekulách DNA rovnaký, molekuly mtDNA nachádzajúce sa v organele nie sú rovnako dlhé (obrázek a obrázek).
„Záhadným“ znakom mitochondriálnych telomér tohto typu je nekompletná repetícia na konci molekuly a relatívne dlhé (110 nukleotidov u Candida parapsilosis) prečnievajúce vlákno na konci 5´. Pripomíname, že jadrové teloméry sú zakončené prečnievaním na konci 3´, ktoré je substrátom pre telomerázu. Preto sa hneď podsúva niekoľko otázok. Prečo DNA-polymeráza nedosyntetizuje medzeru dlhú 110 nukleotidov na konci molekuly? Ako je prečnievajúce vlákno na konci 5´ utvorené? Ako je stabilizované? A ako sú vôbec mitochondriálne teloméry tohto typu replikované?
Jednou z možných odpovedí na tieto otázky je izolácia proteínov špecificky interagujúcich so sekvenciami v telomerickej oblasti (spomeňme si, ako izolácia cicavčích proteínov TRF1 a TRF2 pomohla identifikovať tslučky cicavčích telomér). Nedávno sa podarilo izolovať proteín mtTBP (mitochondrial Telomere Binding Protein), ktorý v laboratórnych podmienkach prednostne viaže telomerické sekvencie a chráni ich pred atakom enzýmov modifikujúcich DNA, čo poukazuje na jeho možnú úlohu v ochrane koncov mtDNA (capping funkcia). Elektrónovomikroskopická analýza ukázala, že mtTBP je v bunkách Candida parapsilosis asociovaný s mitochondriálnymi telomérami, čím sa potvrdil predpoklad, že mtTBP je špecifickou zložkou mitochondriálneho telomerického chromatínu. Analýza sekvencie aminokyselín mtTBP ukázala, že patrí do skupiny tzv. proteínov SSB (single-stranded DNA binding), ktoré sa podieľajú na replikácii, rekombinácii a reparácii mtDNA. Na rozdiel od mtTBP však nemajú „obľúbenú“ DNA sekvenciu, na ktorú by sa viazali s vyššou afinitou. MtTBP sa tak stal popri proteíne Rap1 Saccharomyces cerevisiae ďalším príkladom promiskuitného DNA-väzobného proteínu, ktorý teloméry „využívajú“ na špecifické funkcie. Pravdepodobný scenár evolučnej adaptácie mtTBP by sme mohli napísať v duchu spomenutej jacobovskej predstavy evolučného „kutilstva“, podľa ktorej sa na riešenie nových evolučných problémov využívajú existujúce bunkové prostriedky. Tým novým problémom v prípade Candida parapsilosis bola linearizácia mtDNA a tým existujúcim prostriedkom bol mitochondriálny proteín SSB, ktorý si pribral ďalšiu úlohu.
Najnovšie výsledky štúdia replikácie mtDNA Candida parapsilosis naznačujú, že ďalšími komponentmi štandardného mitochondriálneho replikačného aparátu, ktoré sa využili na replikáciu telomér, sú rekombinačné proteíny. Technikami dvojrozmernej agarózovej elektroforézy a elektrónovej mikroskopie sa v mitochondriách tejto kvasinky identifikovali kružnicové molekuly DNA, ktoré sú zložené výhradne z telomerických tandemových repetícií. Ich veľkosť možno vyjadriť vzorcom n × 738 bp, kde sa hodnota n pohybuje medzi 1–12. Kružnicové mtDNA telomér sú podobné APB-štruktúram, ktoré R. Reddel pozoroval v nádorových bunkových líniách bez aktívnej telomerázy. Štúdium mechanizmu ich tvorby a účasti na replikácii mitochondriálnych telomér preto môže mať význam na pochopenie alternatívnych mechanizmov replikácie jadrových telomér. Posledné tvrdenie sa dá zovšeobecniť: každá stratégia replikácie telomér predstavuje unikátne riešenie a nádorové bunky, ktoré sa vzopreli Hayflickovmu limitu alebo chemoterapeutikám ochromujúcim aktivitu telomerázy, ho môžu „objaviť“.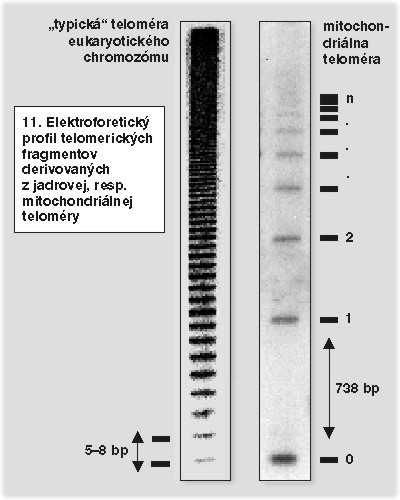
Okrem toho predstavuje časť organizmov s lineárnou mtDNA klinicky zaujímavú skupinu. Mnohé z nich sú totiž primárnymi či potenciálnymi ľudskými patogénmi. Keďže ľudská mtDNA je kružnicová, môžeme si predstaviť liečivá, ktoré špecificky inhibujú replikáciu mitochondriálnych telomér patogéna bez vedľajších účinkov na ľudské bunky. Teloméry sa tak popri gerontológii a onkológii môžu stať zaujímavým terčom aj pri liečbe niektorých infekčných ochorení.
Záverečné poučenie
Učebnice musia nevyhnutne veci zjednodušovať. Preto v nich o telomérach nenájdeme viac ako niekoľko strán textu a obrázkov, ktoré opisujú ich replikáciu prostredníctvom telomerázy. Sú to nepochybne elegantné schémy znázorňujúce fascinujúci enzým. Bunka si však nepotrpí na eleganciu a má k dispozícii ďalšie záložné mechanizmy replikácie telomér. V súčasnosti poznáme okolo tucta rôznych stratégií replikácie koncov lineárnych molekúl DNA a pravdepodobne ich bude ešte viac. Každá z nich si zasluhuje pozornosť. Nielen preto, že má potenciálny klinický význam, ale aj (a snáď hlavne) preto, že ich odhaľovanie je vzrušujúcim intelektuálnym zážitkom. A v tom je aj pointa nášho príbehu. 1)Literatura
www.genlink.wustl.edu/teldb/index.htmlBlackburn E. H., Greider C. W.: Telomeres. Cold Spring Harbor Laboratory Press, 1995
Greider C. W., Blackburn E. H.: Telomeres, Telomerase and Cancer. Sci. Amer. 274, 80–85, 1996
McEachern M. J., Krauskopf A., Blackburn E. H.: Telomeres and their control. Annu. Rev. Genet. 34, 331–358, 2000
Nosek J., Tomáška Ľ., Suyama Y., Fukuhara H., Kováč L.: Linear mitochondrial genomes: 30 years down the line. Trends Genet. 14, 181–188, 1998
Poznámky
Ke stažení
 Článek ve formátu PDF [731,33 kB]
Článek ve formátu PDF [731,33 kB]
O autorech
Jozef Nosek
Ľubomír Tomáška
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
