










Srůstání organizmů
Sekvencování celých genomů dalo nový význam otázce, zda informace v buněčné DNA bezezbytku popisuje eukaryontní buňku. Vhled získaný studiem evoluce a vývoje mitochondrií i chloroplastů naznačuje, že nikoli. Výstavba těchto organel vyžaduje kupříkladu složité mechanizmy pro přísun proteinů, vznikajících v cytoplazmě, na různá místa v organelách. Tyto mechanizmy přemísťují desítky proteinů v cytoplazmě, v membránách i vnitřních prostorách organel a ke své funkci vyžadují přesnou orientaci ve vnitrobuněčných a transmembránových vztazích. Současné poznatky jednoznačně svědčí o tom, že zmíněné dráhy nejsou schopny se uspořádat, jestliže neexistuje jejich předchozí kopie (výstavba je závislá na vzoru). Proto je každý protein potřebný pro přísun jiných proteinů do mitochondrií kvasinky životně důležitý, a to za všech podmínek. Totéž bude pravděpodobně platit pro chloroplasty, a zřejmě i pro organely, které nemají vlastní genom, jako jsou peroxizomy, Golgiho aparát nebo endoplazmatické retikulum.
Závažná role membrán se předpokládala už dříve, ale dnes máme informace k jejímu pochopení na molekulární úrovni. Je pro nás naléhavou výzvou, abychom se pokusili určit, jak velká část našeho genetického dědictví je zakódována ve strukturách našich buněčných organel. 1)
G. Schatz
O dvou typech buněčných organel - mitochondriích a chloroplastech - se předpokládá, že v evoluci vzešly z bakterií, které začaly symbioticky žít uvnitř eukaryontních buněk. S bakteriemi sdílejí především tyto vlastnosti:
- Nikdy nevznikají de novo - množí se pouze dělením mateřské organely.
- Mají genetický aparát bakteriálního typu, odlišný od struktur a procesů v jádře a cytoplazmě buňky. Mitochondrie vzešly ze skupiny alfa-proteobakterií, chloroplasty ze sinic.
- Některé membránové lipidy těchto organel (např. kardiolipin) najdeme v bakteriích, ale nikoli v ostatních membránách téže buňky.
- Srovnáme-li pořadí aminokyselin v proteinech vykonávajících stejnou funkci v organele a v cytoplazmě, obvykle se ukáže, že jde o produkty dvou různých genů. Organelární protein je svými parametry víc příbuzný proteinům volně žijících bakterií než proteinům z jiných částí vlastní buňky.
- Obě organely jsou sídlem některých funkcí, které jsou jinak přítomny jen v bakteriích; nejdůležitější z nich jsou dýchání, fotosyntéza a syntéza adenozintrifosfátu (ATP) pomocí membránového komplexu F0F1-ATPsyntetázy (viz Vesmír 77, 23, 1998/1).
Pro pochopení, jak v evoluci k symbiózám došlo, jsou důležité endosymbiózy řady organizmů s bakteriemi. Lze popsat mnoho závislostí, od parazitů (např. Salmonella) až po velmi těsné svazky obou typů buněk, takové, že ani jeden z organizmů není schopen samostatně žít bez partnera.
Jak se organely zapojily do života buňky
Mitochondrie i chloroplasty obsahují řádově 1000 proteinů. Jejich genom by proto měl obsahovat právě tolik genů. Avšak genom chloroplastů obsahuje jen 100200 genů, a genom mitochondrií dokonce jen několik desítek. Z tohoto počtu jen část představují geny pro proteiny (zbytek jsou geny pro RNA).Počet proteinů syntetizovaných v mitochondriích kolísá od 62 u bičíkovce Reclinomonas americana přes 37 u játrovky Marchantia, 13 u savců až k pouhým 3 u původce malárie Plasmodium falciparum.
Jelikož volně žijící bakterie mají asi 3000 genů, znamená to, že z genomu původních symbiontů jich většina během proměny v organelu zmizela. Část se jich nepochybně ztratila - například geny kódující některé proteiny buněčných povrchů a řadu metabolických drah.
Ty proteiny, které v organelách jsou, a přitom tam svůj gen nemají, musí mít své geny v jádře, jsou syntetizovány v cytoplazmě, a posléze přesunuty do organely. Může jít samozřejmě i o produkty genů, které byly v jádře odjakživa. Jelikož však řada organelárních proteinů vykazuje nepochybné bakteriální signatury, jejich geny byly do jádra patrně přesunuty z původního bakteriálního symbionta. Přesun byl spojen s nutnými změnami v oblasti DNA před genem a za ním - tyto regulační oblasti jsou v jádře jiné než v bakteriálním genomu.
Nevíme, jak se geny do jádra dostaly, ale nejspíš se jich většina přesunula hned v počátcích soužití, a potom - u různých skupin organizmů různě - je tu a tam následovaly geny další. Tak se stalo, že z různých proteinů syntetizovaných v mitochondriích jen dva jsou tam syntetizovány vždy (cytochrom b a jedna podjednotka cytochromoxidázy). Ostatní tvoří proměnlivou množinu, podle toho, co zrovna u dané vývojové linie v organele přežilo. Řád v tom není a neví se, proč vůbec se mitochondrie namáhají s udržením genetického aparátu a syntézou těch několika proteinů. Jde o proteiny vysoce hydrofobní, a tak se původně myslelo, že buňka nebyla schopna vyřešit jejich transport přes vodní prostředí cytoplazmy. Předpoklad sice nebyl zavržen úplně, ale omezil se nakonec jen na zmíněné dva proteiny.
A s transportem začínají problémy. Protein je sice syntetizován jako lineární řetězec aminokyselin, ale ihned se balí do trojrozměrné struktury, která sama o sobě do membrán a vnitřních prostor organel vstupovat nemůže. Buňka tedy musela vyvinout složitý systém pro transport proteinů do organel, zahrnující zejména soustavu chaperoninů (viz Vesmír 77, 15, 1998/1) a složitých membránových pórů. Důležité z tohoto hlediska jsou dvě skutečnosti:
- Původní bakteriální gen kromě zmíněných regulačních sekvencí musel získat novou část kódující signální konec preproteinu. Schatz ve své přednášce na citované konferenci zdůraznil, že to nemusel být zas tak velký problém: Tyto signální konce, které plní funkci jakéhosi směrového čísla, mají šroubovicovou strukturu, a té lze dosáhnout z velkého množství nukleotidových sekvencí.
- Komponenty přívodního systému musely být do organely dopraveny popsaným způsobem! Opět jsme u známého cyklu slepice - vejce, a právě na tuto skutečnost upozorňuje Schatz: organely ani jejich přívodní systémy nemohou vznikat de novo, protože k jejich výstavbě je potřeba informace o předchozím vývoji. Vskutku, jediné smrtící mutace genů pro mitochondriální proteiny se (u kvasinek) týkají genů pro proteiny, které jsou součástí přívodní dráhy.
Další evoluční souvislosti
Jedna skupina eukaryontních organizmů bez mitochondrií (Archaeozoa), protisti ze skupiny Microsporidia a Diplomonadina, jako by žádné symbiotické organely neměla. Tyto organizmy byly proto dlouho považovány za prastaré skupiny eukaryont, které se odštěpily ještě před nástupem endosymbiózy. Teorie byla nedávno zpochybněna, když i u těchto organizmů byly nalezeny geny kódující u ostatních protein sídlící v mitochondriích. Přítomnost takového genu v buňce bez mitochondrií může být vysvětlena tak, že kdysi skupina mitochondrie měla. Jejich geny se přenesly do jádra, načež organely zanikly. Z kočky zbyl jen škleb.
Jiná skupina archezoí (Parabasalia) má hydrogenozom, zvláštní organelu, která zpracovává produkty anaerobního kvašení a produkuje vodík. Organela neobsahuje žádnou DNA. Podobně jako mitochondrie a chloroplasty nevzniká de novo, ale jen dělením organel dosavadních, což vzbudilo podezření, že vznikla endosymbioticky, např. z anaerobních bakterií typu Clostridium. Jedinou slabinou této hypotézy je skutečnost, že se hydrogenozomy vyskytují i u několika skupin nálevníků a hub, organizmů, jejichž nejbližší příbuzní mitochondrie mají. Mnohonásobná endosymbióza se přecejen nejevila jako příliš věrohodná, a tak to vypadá, že i v tomto případě jde o pozměněné mitochondrie. V prostředí bez kyslíku si mitochondrie mohly dovolit ztratit svůj genetický aparát a spoustu dalších funkcí. Nějaké funkce potom naopak získaly a vznikl z nich hydrogenozom. Ve prospěch této verze mluví i nález Hsp70 mitochondriálního typu v hydrogenozomech, a dále to, že pokud genem pro hydrogenozomální protein transformujeme organizmus s mitochondriemi, bude syntetizovaný cizí protein přenášen právě do mitochondrií. A tak by se mohlo ukázat, že všechny bezmitochondriální organizmy jsou nakonec jen odvozeninami a žádná skutečná Archezoa už dnes nežijí.
Poznámky
Transport do mitochondrií
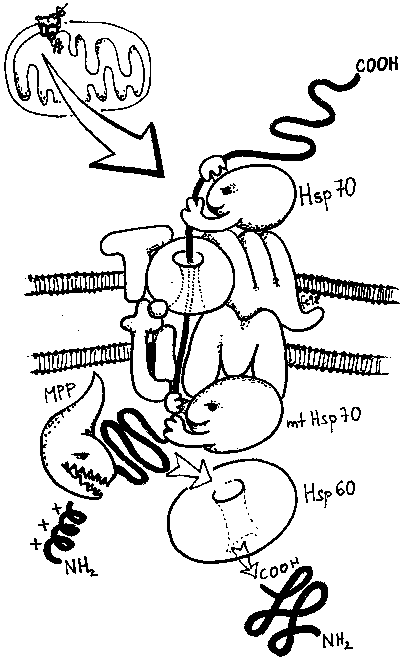
- Protein (přesněji preprotein) syntetizovaný v cytoplazmě musí nést signální sekvenci, směrové číslo, úsek na molekule, který umožňuje jeho rozpoznání jako proteinu určeného do mitochondrií. Je jím krátký úsek na začátku řetězce, uspořádaný do šroubovice tak, že jedna její strana je po délce hydrofobní, druhá kladně nabitá.
- Takto označené proteiny jsou uchopeny systémem chaperoninů (Hsp) a dalších pomocných proteinů a dopraveny k vnější mitochondriální membráně, kde se systém naváže na multiproteinový pór označovaný jako TOM (Translocase of the Outer Membrane). Protože molekula ve tvaru klubíčka se přes TOM nedostane, je nutno ji rozbalit a jako jednorozměrný řetězec soukat přes vnější mitochondriální membránu.
- Jestliže je protein určen pro mitochondriální matrix, musí překonat ještě vnitřní mitochondriální membránu. V té se nachází pórový komplex TIM (Translocase of the Inner Membrane, složením naprosto odlišný od TOM). Představa je taková, že obě membrány k sobě přilnou a protein prochází komplexem TOM/TIM. Zavedení do TIM vyžaduje přítomnost membránového potenciálu na vnitřní membráně, tj. mitochondrie musí být energeticky nabité.
- Z vnitřní strany je preprotein opět uchopen chaperoninem (Hsp70 bakteriálního, tj. i mitochondriálního typu), pomocí proteinázy (MPP) je odštěpena signální sekvence, a působením dalšího chaperoninu získá konečné uspořádání.
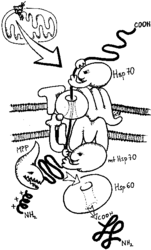 Pro proteiny určené do obou membrán a mezimembránového prostoru existují variace na popsané téma. Proteiny vnější membrány nemají signální sekvenci, vážou se sice na TOM, ale nepřesouvají se a vnoří se do membrány. Proteiny určené do mezimembránového prostoru a vnitřní membrány mohou, ale nemusí mít signální koncovou sekvenci (signál je na jiné části molekuly), a komplex TOM je složen z jiných komponent. Do vnitřní membrány jsou proteiny zabudovány systémem TIM, který je sice podobný, ale nezávislý na výše popsaném.
Pro proteiny určené do obou membrán a mezimembránového prostoru existují variace na popsané téma. Proteiny vnější membrány nemají signální sekvenci, vážou se sice na TOM, ale nepřesouvají se a vnoří se do membrány. Proteiny určené do mezimembránového prostoru a vnitřní membrány mohou, ale nemusí mít signální koncovou sekvenci (signál je na jiné části molekuly), a komplex TOM je složen z jiných komponent. Do vnitřní membrány jsou proteiny zabudovány systémem TIM, který je sice podobný, ale nezávislý na výše popsaném.
Transport do chloroplastů
U chloroplastů přistupuje ještě třetí systém membrán tylakoidy. Věnujme pozornost jen vnější a vnitřní membráně, které jsou analogické mitochondriálním. Situace je na první pohled shodná s mitochondriemi: signální sekvence, membránové póry (TOC a TIC), chaperoniny. Komponenty celého systému však nejsou s mitochondriálními homologní obě dráhy se liší snad ve všech detailech. Chloroplasty vstoupily do symbiotického vztahu později než mitochondrie, evidentně však odmítly převzít od svých předchůdců jejich mechanizmy přenosu a vypracovaly si své vlastní.
Ke stažení
 Článek ve formátu PDF [59,11 kB]
Článek ve formátu PDF [59,11 kB]
O autorovi
Anton Markoš
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
