









Vázaný a nevázaný život lišejníků
K devadesátým narozeninám prof. Zdeňka Černohorského
Jedna ze základních vlastností života, která fascinuje odborníky i laiky, je adaptace na měnící se prostředí (Vesmír 66, 216, 1987/4). Zejména přizpůsobení velmi nepříznivým podmínkám často přesahuje lidskou představivost. Mnohé organizmy objevíme i na velmi nehostinných místech, na nichž bychom již život neočekávali. Týká se to například polárních oblastí, kde najdeme některé zástupce řas, sinic i lišejníků jako endolitické formy života. Žijí nikoli na povrchu kamenů a valounů, ale v hloubce několika milimetrů. V porézním kameni, např. pískovci, osídlují mikroskopické puklinky nebo dutiny.
Lišejníky jsou jednak příkladem života v drsných podmínkách, jednak učebnicovým vzorem symbiózy. Skutečný vztah zúčastněných partnerů je však často daleko mnohotvárnější, než se všeobecně ví, a to nejen z hlediska jejich počtu, ale i charakteru vzájemného působení. Dokonce má i více tváří, které mají ke kooperativnímu ideálu symbiózy daleko.
Strategie heterotrofů a lichenizace
Houby, dnes řazené do samostatné říše, nebo dokonce i do několika říší, jsou klasickým příkladem heterotrofních organizmů. Oproti autotrofním organizmům, které si organické sloučeniny vytvářejí fotosyntézou, mají komplikovanější způsob života, neboť jsou odkázány na jiný zdroj organických látek. Strategií umožňujících přístup k organickým látkám je v podstatě několik a houby všechny tyto způsoby využívají:- saprofytizmus – pomalý přístup umožňující zpracování těl již odumřelých organizmů nebo výkalů;
- predace – lov kořisti, speciálními mechanizmy k lovu jsou vybaveny i některé houby;
- parazitizmus – příživnictví, život na úkor jiného organizmu; parazit je s ním po určitou dobu svým životním cyklem těsně svázán;
- symbióza – soužití, které přináší výhody pro všechny partnery, jde však o způsob nejednoznačný a komplikovaný, jak se vzápětí ukáže.
Lišejník není organizmus, ale asociace
Zmíněné strategie nejsou vázány na jednotlivé vývojové linie, mohou se však velice lišit i u příbuzných skupin, ba dokonce se mohou střídat během života jedince (ontogeneze). A navíc – v průběhu fylogeneze se stejný typ strategie vyskytl několikrát. Lichenizace tedy nepředstavuje jednotný princip – jak lichenizované, tak nelichenizované zástupce můžeme najít i mezi zástupci blízce příbuznými, dokonce v rámci jednoho rodu. Lišejníky (Lichenes) proto nejsou skupinou taxonomickou, nýbrž ekologickou a je správnější o nich mluvit jako o lichenizovaných houbách. Přísně vzato, lišejník není organizmus, ale asociace. Tato skutečnost má i praktické důsledky: vědecké jméno lišejníku neoznačuje celou asociaci (tedy lišejník), váže se pouze na lichenizovanou houbu, zatímco fotosyntetizující partner má své vlastní jméno rodové i druhové.Lichenizace není u hub jediným případem symbiózy. Jiným, rovněž dosti rozšířeným a pro oba partnery také evolučně přínosným způsobem je mykorhiza tedy soužití hub s kořeny cévnatých rostlin, a to jak stromů, tak bylin. Z celkového počtu druhů hub (téměř 65 000) tvoří lichenizované houby 21 % a mykorhizní asi 8 %.
Lišejník není jen houba a řasa
Lišejníky jsou obecně známým příkladem symbiotického soužití houby s řasou. Ve skutečnosti se však jako autotrofní fotosyntetizující partner – fotobiont – mohou vyskytovat též sinice (asi u 8 % druhů lišejníků). Houbový partner – mykobiont – bývá nejčastěji zástupce hub vřeckovýtrusých (Ascomycetes), v několika případech stopkovýtrusých (Basidiomycetes). Dnes běžně známý fakt, že lišejník je symbiotický organizmus složený z houby a řasy, však čekal na odhalení až do poloviny minulého století, a tento objev nebyl nijak lapidární. Vědělo se, že stélka lišejníků obsahuje nazelenalé kulovité útvary, avšak ty se považovaly za orgány nepohlavního rozmnožování a označovaly se jako gonidie (pojmenovány analogicky ke konidiím, nepohlavním rozmnožovacím částicím hub).  Teprve r. 1867 švýcarský rostlinný anatom Simon Schwendener odhalil a správně pojmenoval skutečnou, tedy podvojnou povahu lišejníků (formuloval dualistickou hypotézu). Jeho poznatek byl vpravdě revoluční a není bez zajímavosti, že největší lichenologové té doby ji nepřijali (např. W. Nylander, G. Koerber, A. Krempelhuber) a boje proti ní se nevzdali po celý svůj život. Plamínek svatého boje proti kacířství hořel kuriózně téměř sto let a práce popírající dualistickou povahu lišejníků byla publikována ještě r. 1953.
Teprve r. 1867 švýcarský rostlinný anatom Simon Schwendener odhalil a správně pojmenoval skutečnou, tedy podvojnou povahu lišejníků (formuloval dualistickou hypotézu). Jeho poznatek byl vpravdě revoluční a není bez zajímavosti, že největší lichenologové té doby ji nepřijali (např. W. Nylander, G. Koerber, A. Krempelhuber) a boje proti ní se nevzdali po celý svůj život. Plamínek svatého boje proti kacířství hořel kuriózně téměř sto let a práce popírající dualistickou povahu lišejníků byla publikována ještě r. 1953. 

Vzájemné vztahy
Vzájemný vztah dvou partnerů může být dvojí:- antagonistický – mykobiont může působit fotobiontu újmu, a to buď lokálně omezenou nekrózu, nebo i jeho odumření.
- mutualistický – vytváří se stabilní asociace bez zjevného poškození fotobionta. Mutualistický typ symbiózy je přínosem pro obě zúčastněné složky.
Takto striktní dělení obou typů soužití je však dáno jen lidským pohledem. Kromě toho lze ještě připomenout komenzalistické soužití, v němž sice jeden partner získává větší prospěch, avšak nikoli za cenu újmy druhého partnera. Navíc vzájemný vztah organizmů může za různých podmínek či okolností procházet proměnami.
U některých lišejníků (asi 500 druhů) se současně mohou vyskytovat dva různí fotobionti, vedle řasy je druhým nejčastějším fotobiontem sinice. Většinou je jeden fotobiont základní a dominující. Tvoří samostatnou vrstvu v rámci lišejníkové stélky, zatímco druhý je izolován do jakýchsi ostrůvků, a to buď uvnitř stélky, nebo častěji v tvarově a barevně nápadných morfologických útvarech na stélce – cefalodiích. Podobně se může počet fotobiontů rozšířit až na tři, kdy se vytvářejí různá cefalodia obsahující rozdílné fotobionty.
Také mykobiontů může být více, a to například tehdy, když je lišejník napaden houbou. Ta však nemusí na hostitele působit vždy vyloženě destruktivně, a proto druhého houbového partnera v takovém případě označujeme jako parasymbionta a příslušný vztah jako komenzalizmus. Houby vázané svým výskytem na lišejníky se označují jako lichenikolní houby a jejich celkový počet se zatím udává kolem 1000 druhů. Parasymbiont získává organické látky od fotobionta, fyziologicky se tedy tyto houby od sebe neliší.
Vzájemný vztah partnerů může být komplikovaně provázán a je lépe ho označit jako asociaci. Na jednom lišejníku se může vyskytovat více parazitů či parasymbiontů, a proto může být vyšší i počet mykobiontů. Další modifikací tohoto typu soužití jsou případy, kdy mykobiont parazituje na existující asociaci, postupně eliminuje původního mykobionta, a posléze vytváří s fotobiontem vlastní mutualistickou stélku (např. Diploschistes muscorum). 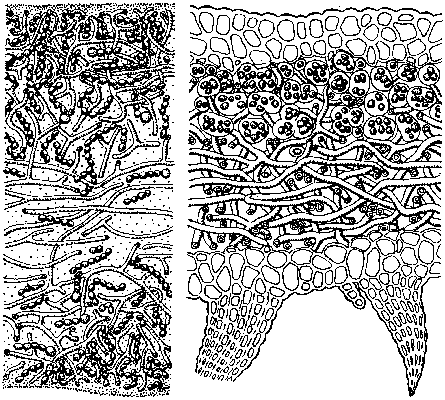
Známe také lišejníky, které alespoň v počátečním stadiu vývinu potřebují ke svému úspěšnému růstu hostitelský lišejník. Na rozdíl od předchozích případů však mají vlastního fotobionta. Tyto lichenikolní lišejníky představují asociaci dvou mykobiontů a dvou fotobiontů. Počet jejich druhů je odhadován na 200 až 300.
Kromě toho existují i mechanické hybridy (pravé chiméry), které vznikly fúzí mycelií nebo mladých stélek pocházejících z několika rozmnožovacích částic stejných či různých druhů, popřípadě dokonce též různých rodů.
Morfologické hrátky lišejníků
Ani základní postulát lichenologie, podle nějž se všechny lišejníky od sebe odlišují tím, že jsou tvořeny jinou houbou, neplatí bez výjimky. Velmi pozoruhodné totiž je, že stejná houba může tvořit odlišné asociace s různými symbiotickými partnery – tedy jedna houba vytváří různé lišejníky. Nejnápadnější rozdíly jsou při účasti zcela odlišných fotobiontů – řas či sinic. Takové stélky mohou mít jinou barevnost nebo odlišný celkový tvar, mohou růst pohromadě nebo být na sobě zcela nezávislé.Nejznámějšími příklady jsou dobře vyvinutá cefalodia, která jsou schopna samostatné existence; tak je tomu např. u hávnatky bradavičnaté (Peltigera aphthosa). Za normálních okolností tvoří tato hávnatka zelenou lupenitou stélku obsahující zelenou řasu jako fotobionta. Na stélce se vytvářejí cefalodia ve tvaru tmavých bradavčitých útvarů, která obsahují sinici jednořadku (Nostoc). Za určitých ekologických podmínek se vytvářejí cefalodia ve tvaru drobných lalůčků, které mohou nést další lalůčky se zelenou řasou. Vznikají tak mozaikovité (chiméroidní) stélky: základní stélka se zelenou řasou jako fotobiontem, cefalodium se sinicí a na něm opět stélka se zelenou řasou. Nejde však o chiméry, neboť stélky jsou tvořeny geneticky uniformní houbou. Navíc ještě může osamostatněné cefalodium vytvořit stélku tvarově shodnou s mateřskou stélkou, ale inverzní: Tmavá stélka obsahující sinici nese na povrchu drobné zelené lalůčky obsahující řasu, na lalůčcích opět mohou růst tmavá cefalodia. Takové případy, kdy jedna lichenizovaná houba vytváří s různými fotobionty různé stélky, se označují jako fototypy nebo fotomorfy.
Ještě podivnější situace nastávají, když obě fotomorfy mají zcela odlišný vzhled a tvoří buď lupenitou, nebo keříčkovitou stélku, např. u některých druhů laločníku (Sticta). Není proto divu, že nesly samostatná jména. Tyto případy také ilustrují to, že fotobiont má někdy zásadní význam pro tvar stélky, přestože v ní má výrazně menšinový podíl a přestože ji stavebně určuje mykobiont.
Aby to bylo ještě zamotanější, jsou známy i případy, kdy z téhož lišejníku byli izolováni různí fotobionti. Mykobiont tedy v těchto případech není specifický vůči určitému fotobiontu a může vytvořit shodnou stélku i s jinou řasou.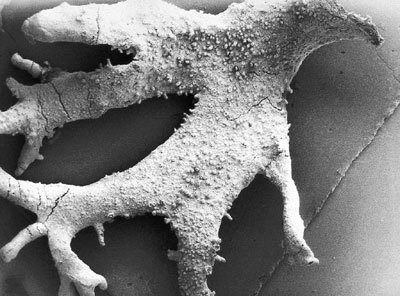
Kde se vzaly lišejníky
Symbióza je natolik složitý vztah provázaný mnoha vzájemnými vazbami, že nelze očekávat jeho vznik krátkou cestou a přímo. Velice pravděpodobně se vyvinul z parazitizmu koevolucí – dlouhým a vzájemným vývojem obou partnerů, jehož počátky jsou velmi staré. Předpokládá se, že většina lichenizovaných čeledí a rodů se vyvinula na rozhraní prvohor a druhohor (před 190–280 miliony let). Možné počátky lichenizace se na základě ne zcela jasného paleontologického nálezu dokonce kladou až do prekambria (před více než 2 miliardami let).
Jeden z možných modelů vzniku lichenizace naznačují mořské řasy Ascophyllum nodosum a Pelvetia canaliculata, hostící houbu Mycosphaerella ascophylli. Ta tvoří plodničky ve stélce řas, které se projevují jako drobné černé skvrny. Přitom se ovšem nijak nezdá, že by houba hostitelské řase škodila. Naopak, nalézt „nenapadenou“ řasu, tedy bez této houby, je daleko obtížnější, a dokonce jsou pochyby, zda vůbec řasa neinfikovaná houbou může v přirozeném prostředí přežívat. Snad je toto soužití jedním z možných stadií vývoje symbiotického vztahu. Do určité míry má charakter lichenizace, nevytváří se však specifická stélka a řasa si zachovává svůj charakter – proto je označován jako mykofykobióza. Podobné vztahy nacházíme i u dalších hub asociovaných s řasami a tento typ soužití není omezen jen na vodní prostředí. Porosty zelených řas typu zrněnky (Desmococcus vulgaris) na borce stromů jsou velmi často asociovány s myceliem hub.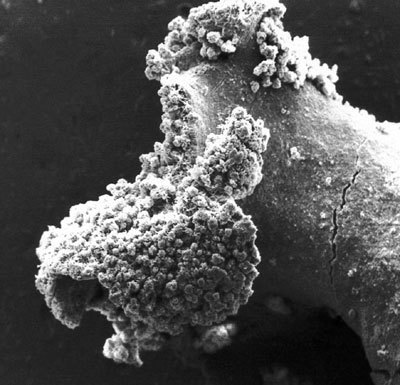
Přestože vznik lišejníkové stélky byl evolučně velkým přínosem pro obě zúčastněné složky, přece jen má houba jako heterotrofní složka z asociace ustanovené lichenizací větší prospěch. Fotosyntetický partner je v područí houby. Ve skutečnosti lichenizace není „opravdovou“ a nezištnou symbiózou, ale v podstatě jde o kontrolovaný parazitizmus. Míra této kontroly je závislá na délce společného soužití a vybalancování tohoto vztahu je tedy úměrná evoluci.
Stélka lišejníků
Lichenizace dává vznik nové entitě – stélce. Ta může být různá: od primitivního kontaktu a promísení téměř nezměněných těl jednotlivých partnerů až po vysoce rozčleněnou stélku se specifickými orgány (Vesmír 53, 69, 1974/3). Jednoduchou stavbu má stélka homeomerická, v níž je fotobiont náhodně rozptýlen. Pro vnitřní stavbu většiny lišejníků je typická vrstevnatá heteromerická stélka, kde fotobiont tvoří samostatnou vrstvu. Ta je kryta pletivem mykobionta – svrchní kůrou. Jednotlivé buňky houbových vláken k sobě těsně přiléhají a mají ztluštělé buněčné stěny, neboť tato vrstva má především ochrannou funkci. Pod vrstvou fotobionta je dřeň, která je opět tvořena pouze mykobiontem; tentokrát jde o řídké pletivo, jehož houbová vlákna jsou propletena do vatovité struktury s množstvím mezibuněčných prostor, sloužících především jako zásobárna vody. Pod dřeňovou vrstvou bývá např. u většiny lupenitých lišejníků vyvinuta spodní kůra, korovité lišejníky většinou k substrátu přirůstají přímo dření.Stélky lišejníků jsou tvarově neobyčejně rozmanité: od velmi drobných, téměř neviditelných zástupců po několikametrovou provazovku (Usnea longissima), od jednoduché korovité stélky, tvořící tenké povlaky na různých substrátech, až po velmi členitou stélku nepravidelných, bizarních tvarů, s níž se setkáváme u stélek lupenitých a keříčkovitých. Proto také lišejníky poutají pozornost jak vnímavých návštěvníků přírody, tak umělců (např. M. Escher použil v litografii Vodopád lišejníky v „nadživotní velikosti“).
Rozmnožování lišejníků
Velkým evolučním přínosem lichenizace je vytváření specifických struktur, které nenajdeme u žádných jiných organizmů. Týká se to především vegetativního rozmnožování.Mykobiont sám většinou vytváří orgány pohlavního i nepohlavního rozmnožování, které se příliš neliší od orgánů nelichenizovaných hub. I když se však fotobiont podílí na stavbě plodniček, vlastního rozmnožování výtrusy se zúčastní pouze mykobiont. Jedinou výjimkou jsou rody Endocarpon a Staurothele, které mají hymeniální řasy. Jsou to zmenšené buňky fotobionta vyskytující se v hymeniu (výtrusorodém roušku) plodnic a při uvolňování výtrusů z vřecek se mohou šířit oba partneři zároveň, okamžitě nastolit symbiotický vztah a vytvořit stélku.
Velkého rozvoje však u lišejníků doznalo rozmnožování vegetativní, které právě umožňuje buď přímo šíření fragmentů stélky, nebo alespoň současné šíření obou partnerů. Specifickými orgány vegetativního rozmnožování u lichenizovaných hub jsou především velmi běžně rozšířené izidie a soredie. Izidie jsou bradavčité útvary na povrchu stélky různého tvaru (pro jednotlivé druhy jsou však charakteristické). Jsou anatomicky rozlišeny a jejich vnitřní stavba odpovídá heteromerické stélce. Lišejníky se šíří ulomením izidií, z nichž regeneruje stélka; transportu se účastní voda či vítr. Soredie jsou oproti tomu velmi drobné (pod 1 mm) kulovité chomáčky houbových vláken, která obklopují shluk několika buněk fotobionta. Nemají tak diferencovanou stavbu jako izidie, zato jsou drobnější a lehčí, což umožňuje šíření na větší vzdálenosti – opět u obou symbiotických partnerů zároveň. Soredie vznikají ve vrstvě fotobionta v sorálu, což bývá ohraničený útvar na vnějším povrchu stélky. Sorály nejsou kryté kůrou a vizuálně se projevují jako lokální zdroj prášku soredií, často i barevně od stélky odlišného. Rovněž sorály mohou být různého typu, který je pro daný druh charakteristický a slouží jako určovací znak.
Biochemická pestrost
Lišejníky se vyznačují velmi specifickými sekundárními produkty metabolizmu. Někdy se těmto látkám říká lišejníkové kyseliny, avšak toto označení není přesné, neboť jde o velmi rozmanité organické látky. Dnes je u lišejníků známo více než 700 sekundárních produktů metabolizmu, které se zpravidla jinde než v lišejnících nevytvářejí (necelá desetina těchto látek se vyskytuje i u nelichenizovaných hub či u vyšších rostlin). Vesměs tyto látky vytváří mykobiont a jsou ukládány vně buněk pouze v určité části lišejníkové stélky, např. na povrchu houbových vláken v dřeni. Některé látky jsou barevné, ale většina jich je bezbarvých a u různých druhů lišejníků mají různou funkci. Mohou sloužit jako filtr chránící fotobionta před nadměrným ozářením. Největší význam mají v obraně vůči jiným organizmům, u řady z nich byly prokázány antibiotické či antiherbivorní účinky a některé jsou dokonce toxické. Nejznámější je kyselina vulpinová, která je obecně jedovatá, a to nejen pro savce, ale též pro hmyz a měkkýše – velmi překvapivě však nepůsobí na králíky a myši. Lišejníkové látky jsou produktem symbiotického soužití; ani jedna izolovaná složka asociace je zpravidla sama nevytváří. Jejich vznik zřejmě indukovala řasa chránící se tak před agresivním parazitizmem houby. Snad je to jeden z mechanizmů udržujících rovnováhu vzájemného soužití – kontrolovaného parazitizmu.Ekologické hledisko lichenizace
Lišejníky, ač nejsou schopny aktivní regulace příjmu vody (patří mezi poikilohydrické organizmy), jsou ekofyziologicky velmi dobře přizpůsobeny extrémním podmínkám. Najdeme je proto tam, kde nejsou ohrožovány konkurencí – v oblastech s nepříznivými podmínkami pro růst větších a rychleji rostoucích rostlin, zejména na skalách, v pouštích, vysokých horách a polárních oblastech.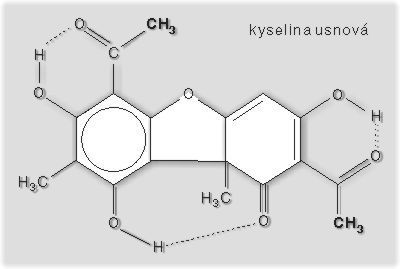 Jejich aktivní projevy života jsou vázány na přítomnost vody ve stélce. Vzhledem k tomu, že lišejníky nemají speciální orgány příjmu vody (přijímají ji celým povrchem těla) a ani nejsou schopny vodu aktivně udržet, je pro jejich život typické neustálé střídání period vlhké a suché stélky. Mohou využívat i vzdušnou vlhkost, déle trvající vlhkost jim však většinou škodí. V klimaticky extrémních podmínkách jsou období vlhké stélky (a tedy aktivního metabolizmu) velmi krátká, a to i v případě, kdy je srážek dostatečné množství, protože zmrzlá voda je pro lišejníky nedostupná. S tím souvisí pomalý růst lišejníků (roční přírůstky se pohybují od zlomků milimetrů až zhruba do centimetru) a dlouhá životnost stélky (až stovky, či dokonce tisíce let).
Jejich aktivní projevy života jsou vázány na přítomnost vody ve stélce. Vzhledem k tomu, že lišejníky nemají speciální orgány příjmu vody (přijímají ji celým povrchem těla) a ani nejsou schopny vodu aktivně udržet, je pro jejich život typické neustálé střídání period vlhké a suché stélky. Mohou využívat i vzdušnou vlhkost, déle trvající vlhkost jim však většinou škodí. V klimaticky extrémních podmínkách jsou období vlhké stélky (a tedy aktivního metabolizmu) velmi krátká, a to i v případě, kdy je srážek dostatečné množství, protože zmrzlá voda je pro lišejníky nedostupná. S tím souvisí pomalý růst lišejníků (roční přírůstky se pohybují od zlomků milimetrů až zhruba do centimetru) a dlouhá životnost stélky (až stovky, či dokonce tisíce let).  Ve fyziologickém spánku odolávají jak nízkým teplotám, tak vyschnutí a přežívají velmi dlouhá období (řádově nejméně měsíce) bez jakékoliv újmy. Lišejníky proto najdeme ve většině biotopů a osídlují i bizarní substráty – kov, sklo či krunýře živých želv a krovky živých brouků. Na 8 % suchozemského povrchu tvoří lišejníky dokonce dominantní složku vegetace.
Ve fyziologickém spánku odolávají jak nízkým teplotám, tak vyschnutí a přežívají velmi dlouhá období (řádově nejméně měsíce) bez jakékoliv újmy. Lišejníky proto najdeme ve většině biotopů a osídlují i bizarní substráty – kov, sklo či krunýře živých želv a krovky živých brouků. Na 8 % suchozemského povrchu tvoří lišejníky dokonce dominantní složku vegetace.
Lišejníkové hádanky a zajímavosti
Stále panují velké nejasnosti v základních věcech. Nevíme, zda jednotliví partneři (hlavně fotobionti) jsou schopni existence i mimo ustanovenou symbiotickou asociaci – zda jde o nezbytnou (obligátní), nebo příležitostnou (fakultativní) symbiózu. Nejběžnějším fotobiontem lišejníků (zhruba u 2/3 z celkového množství druhů) je řasa Trebouxia. Přestože se v literatuře zmiňují nálezy volně žijících buněk či kolonií této řasy, asi půjde jen o ojedinělé a dočasné výskyty (např. buněk uvolněných z mechanicky poškozené stélky). Všeobecně se soudí, že evolučně dlouhým soužitím vznikl vzájemně vyvážený vztah a žádný z partnerů není vzhledem k nastalým proměnám schopen samostatné existence.Okamžitě se však nabízí otázka: Jak se vlastně lišejník rozmnožuje? Kromě zmíněných způsobů vegetativního rozmnožování a hymeniálních řas v plodničkách lišejníků je nutné, aby klíčící spora houby našla odpovídajícího partnera. Jak k tomu však dojde, když se řasa nevyskytuje volně? Údaje o pozorování vzniku stélky v přírodě jsou zatím jen sporé. Jedním ze způsobů je vytváření dočasných asociací s volně žijícími řasami, aniž by vznikla stélka. Dalším způsobem může být parazitizmus na stélce již ustanovené, resp. jejím fotobiontu. 
Pozoruhodný je případ nedávno zaznamenaný u běžného lišejníku nápadné oranžové barvy Xanthoria parietina. Jeho stélka obsahuje fotobionta, který je v přírodě velmi vzácný, takže klíčící výtrusy skoro nemají šanci ho nalézt. Normálně nevytváří orgány vegetativního rozmnožování, přesto je však v přírodě hojný. Vývoj stélky začíná klíčením výtrusu uvolněného z vřecka. Vstoupí do soužití s běžnou volně žijící řasou typu zrněnky, která se ale ve stélce nikdy nevyskytuje. Hyfy mykobionta přerůstají povlak řasy na kůře stromů nebo na kameni a vzniká zelená krusta bez jakékoli diferenciace, takže o stélce ještě nelze hovořit. Tato krusta již konkurencí ale vyloučí možnost růstu jiných druhů. Pokud mykobiont druhu Xanthoria parietina narazí na buňku odpovídajícího fotobionta, okamžitě se začne vyvíjet strukturovaná stélka s drobnými lalůčky a rovněž se začne vytvářet charakteristický oranžový pigment parietin.
Odpovídající řasu je houba schopna získat i tak, že ji ukradne ze soredie nebo přímo ze stélky jiného lišejníku (např. šedě zbarveného terčovníku Physcia tenella). Úspěch a především rozsah ataku je závislý na stáří (stadiu ontogeneze) okrádaného jedince. Jestliže dojde k útoku v počátečním stadiu vývinu, je původní mykobiont potlačen a stélka dostává tvar zloděje. Stane-li se to později, je využita pouze část stélky a vzniká mezirodová chiméra. Hyfy zlodějského mykobionta X. parietina mohou pronikat i do soredií olupovaného lišejníku P. tenella, které obsadí, zbarvují je do oranžova (normálně jsou zbarvené světle) a posléze mohou být rozšiřovány i takto vzniklou stélkou (X. parietina totiž normálně soredie nevytváří). Tento dosud ojedinělý způsob rozmnožování ukazuje na velkou tvárnost a pozoruhodné schopnosti lichenizovaných hub. Je pravděpodobné, že bude v přírodě častější. Nejspíš ale lišejníky ovládají i další typy rozmnožovacích strategií.
Různé tváře lichenizace
Vzhledem k tomu, že lichenizace není jednotnou vývojovou linií a byla založena polyfyleticky, není vztah různých lichenizovaných hub k fotobiontu u všech lišejníků shodný a charakter symbiotického vztahu má mnoho podob. Některé typy fotobiontů se v přírodě vyskytují volně. Ne u všech lišejníků je vztah obou partnerů – řasy a houby – velmi těsný, někdy se ani nevytvoří vysoce diferencovaná stélka nebo může být symbiotický vztah omezen jen na určité ontogenetické stadium. U mnoha zástupců, dříve považovaných za lišejníky, se nakonec ukázalo, že nežijí ve stabilní asociaci s řasou, ale pouze porůstají či prorůstají kolonie řasových buněk vyskytujících se v nižších vrstvách borky dnes se tedy označují jako houby nelichenizované.Průměrné roční přírůstky některých lišejníků (v mm)
korovité druhy
Diploschistes scruposus 0,44 - 3,0
Graphis scripta 0,85
Porpidia albocaerulescens0,70
Rhizocarpon geographicum 0,06 - 0,5
lupenité druhy
Brodoa intestiniformis 1,0
Parmelia caperata 3,45 - 6,98
Parmelia glabratula 1,68 - 1,89
Parmelia sulcata 1,6 - 2,2
Peltigera rufescens 25,0 - 27,0
Umbilicaria deusta 2,3
Umbilicaria proboscidea 0,17
Xanthoria parietina 2,5
keříčkovité druhy
Cladonia arbuscula 4,6
Cladonia rangiferina 2,7 - 6,0
Evernia prunastri 2,0
Ke stažení
 DATA A SOUVISLOSTI - Život lišejníků [316,68 kB]
DATA A SOUVISLOSTI - Život lišejníků [316,68 kB]
O autorovi
Jiří Liška
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
