









Kde se v mozku tvoří paměť?
Hipokampová formace je mozková struktura, která hraje důležitou úlohu v paměťových procesech. Důkazem jsou data získaná při sledování neurologických pacientů s úrazem v této oblasti. Poškození hipokampu nebo jeho léčebné odnětí vede u lidí k anterográdní amnézii – postižený si pak není schopen zapamatovat nová fakta a rozpoznávat nové tváře. Dříve vytvořené paměťové stopy však zůstávají neporušené, protože jsou pravděpodobně uloženy v jiné mozkové struktuře, v kůře. Navíc si postižený uchovává typy paměti s hipokampem nesouvisející, jako procedurální paměť, která je potřebná pro získávání dovedností. Klasickým příkladem je dodnes žijící pacient, kterému v roce 1953 byla při neléčitelné epilepsii zároveň s vnitřními částmi spánkových laloků operačně odňata i velká část hipokampu. Pooperační psychologická vyšetření ukázala, že zatímco rysy osobnosti a inteligence zůstaly nenarušeny, pacient si nebyl schopen vybavit jedinou událost počínaje dnem operace, či poznat lidi, se kterými se po operaci seznámil a byl s nimi v kontaktu.
 U hlodavců je hipokampus zapojen do mechanizmu prostorového učení. Zatímco laboratorní potkan s oboustranně odňatým hipokampem není schopen najít ve vodní nádrži podlážku pod hladinou vody, kontrolní zvíře se rychle naučí plavat přímo k ní. Dalším důkazem byl objev hipokampových „buněk určitého místa“. U laboratorních potkanů je každé nové prostředí zmapováno skupinou neuronů v hipokampu. V rámci této skupiny se pak aktivita jednotlivých neuronů váže k přesně definovanému místu (poli) v prostoru. Tak zvíře získává poznatky o prostředí.
U hlodavců je hipokampus zapojen do mechanizmu prostorového učení. Zatímco laboratorní potkan s oboustranně odňatým hipokampem není schopen najít ve vodní nádrži podlážku pod hladinou vody, kontrolní zvíře se rychle naučí plavat přímo k ní. Dalším důkazem byl objev hipokampových „buněk určitého místa“. U laboratorních potkanů je každé nové prostředí zmapováno skupinou neuronů v hipokampu. V rámci této skupiny se pak aktivita jednotlivých neuronů váže k přesně definovanému místu (poli) v prostoru. Tak zvíře získává poznatky o prostředí.
Hipokampus souvisí s učením
Otázkou zůstává, jaký je buněčný mechanizmus tvorby a uchovávání paměťové stopy – jakým způsobem se informace o podlážce ve vodní nádrži ukládá či jak se vytvoří kognitivní mapa vnějšího prostředí. V posledních desetiletích vzbudila pozornost neurovědců synaptická plasticita. To je schopnost dlouho udržet změnu účinnosti synaptického přenosu mezi dvěma neurony a je vyvolána specifickým podrážděním neuronu ležícího před synapsí (zápojem). Často uváděným příkladem role synaptické plasticity při tvorbě paměti je dlouhotrvající dráždění popsané u mořského měkkýše Aplysia californica. Tato jednoduchá forma učení je vyvolána opakovaným působením bolesti na hlavě nebo ocasu měkkýše, což vede k dlouhodobému obrannému zatažení dýchacího orgánu. Na buněčné úrovni se zesiluje synaptický přenos v rámci několika synapsí mezi senzorickými neurony, interneurony a motorickými neurony. Podobně je tomu u obratlovců, kde další typ synaptické plasticity – dlouhotrvající zesílení synaptického přenosu (long term potentiation, LTP) – je jedním z možných mechanizmů tvorby paměti v hipokampu. Model je zajímavý tím, že synaptická plasticita má rysy, které jsou pro mechanizmus paměti nepostradatelné.Přes intenzivní snahu prokázat souvislost mezi dlouhodobým zesílením synaptického přenosu a pamětí se přímých důkazů stále nedostává. Nejpřesvědčivější jsou klasické studie R. Morrise a kolektivu, kteří ukázali, že prostorové učení je stejně jako dlouhodobé zesílení synaptického přenosu blokováno antagonistou receptoru NMDA. K témuž výsledku nyní dospěly studie na transgenních (geneticky modifikovaných) zvířatech. Rychle se vyvíjející metodiky molekulární biologie umožnily vývoj zvířat, kde modifikace genetické informace je prostorově lokalizována pouze do části hipokampu označované CA1. V jedné z těchto studií byla v CA1 odstraněna základní stavební podjednotka receptoru NMDA. Tato manipulace funkci receptoru NMDA narušila. Tím způsobila nedostatek jak synaptické plasticity, tak prostorového učení, a obměnila „buňky místa“. Nový přístup potvrzuje, že alespoň část hipokampu je nezbytná pro prostorové učení. Navíc tyto experimenty přinášejí první důkazy funkční souvislosti mezi prostorovým učením a „buňkami místa“, a rovněž mezi synaptickou plasticitou a tvorbou polí „buněk místa“.
Typy synaptické plasticity
Popsat v jednom článku všechny formy synaptické plasticity v hipokampové formaci a nesklouznout k pouhému výčtu dnes už objevených mechanizmů je nemožné. Proto se soustředíme na oblast CA1, kde výsledky získané na transgenních zvířatech přinášejí další důkazy o funkčním významu synaptické plasticity pro mechanizmy paměti.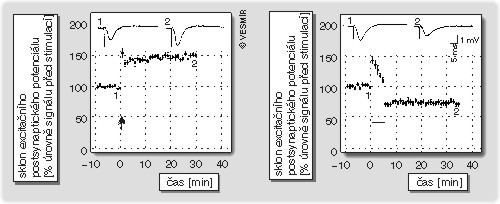
V zásadě existují dva typy synaptické plasticity, dlouhotrvající potenciace (long-term potentiation, LTP) a dlouhotrvající deprese (long-term depression, LTD), které mají na sílu synaptického přenosu opačný účinek: zatímco dlouhotrvající potenciace synaptický přenos zesiluje, dlouhotrvající deprese jej snižuje (obr. obrázek). Zesílení nebo zeslabení synaptického přenosu závisí pouze na frekvenci stimulace. Vysokofrekvenční stimulace (typicky 100 Hz) vede k dlouhotrvající potenciaci, kdežto nízkofrekvenční stimulace (typicky 3 Hz) vede k dlouhotrvající depresi. Naproti tomu asociační plasticita vyžaduje aktivaci dvou vstupů, kde testovací vstup, jehož efektivita má být obměněna, je vzbuzen nízkou frekvencí (typicky 5 Hz) a podmiňovací vstup krátkými opakovanými vysokofrekvenčními pulzy. Zatímco podmiňovací vstup je vždy zesílen, testovací vstup může být buď zesílen, nebo zeslaben. O směru změny na testovacím vstupu rozhoduje pouze časová souvztažnost mezi stimulací testovacího a podmiňovacího vstupu. Aktivace obou vstupů zároveň vede k dlouhotrvající potenciaci, kdežto časový posun mezi nimi vede k dlouhotrvající depresi. Synaptická plasticita splňuje pravidlo, jež ve čtyřicátých letech formulovala D. O. Hebb: Synapse, která spojuje dva neurony, je posílena, pokud jsou obě nervové buňky aktivovány současně. Toto pravidlo lze aplikovat nejen na dlouhodobou potenciaci, ale i na dlouhodobou depresi, a směr změny synaptického přenosu bude pravděpodobně určen mírou aktivace postsynaptického neuronu. Kategorií, která se Hebbovu pravidlu vymyká, je heterosynaptická dlouhotrvající deprese. Tento typ plasticity by se mohl označit jako vedlejší produkt dlouhotrvající potenciace. Po indukci dlouhotrvající potenciace je totiž synaptický přenos na některých synapsích téhož postsynaptického neuronu zeslaben. Jsou to ty synapse, které neobdržely podnět z presynaptické části, ale jejich postsynaptická část byla aktivována díky šíření depolarizace z povzbuzených synapsí. Funkční význam popsaného jevu není znám.
Změny synaptické efektivity vyvolané zesílením i zeslabením přenosu jsou sice dlouhotrvající, nicméně synapse si uchovávají schopnost být dále modifikovány. Například synapse, které jsou posíleny, zůstávají vnímavé jak k další vysokofrekvenční stimulaci (až do stadia, kdy je dlouhotrvající účinek naplněn), tak k stimulaci nízkofrekvenční. Synaptický přenos tak může být zesilován či zeslabován, což přidává systému na přizpůsobivosti a kapacitě pro ukládání informací.
Aktivované receptory
Klíčovou úlohu při zesílení či zeslabení přenosu hraje aktivace receptorů NMDA (excitačních receptorů N-methyl-D-aspartátové kyseliny, patřících do skupiny glutamátových receptorů), protože aplikace antagonistů (opačně působících činitelů) tohoto receptoru blokuje obě formy synaptické plasticity. Je to receptor ionotropní, to znamená, že obsahuje kanál v membráně neuronu a ten se při navázání agonisty (stejně působícího činitele) otevře a umožní přechod iontů přes membránu. Aktivovaný receptor NMDA, který stejně jako další typ ionotropního receptoru AMPA propouští sodík a draslík, je navíc propustný pro vápník a umožňuje jeho vstup do neuronu. Jaký je rozdíl mezi těmito dvěma typy glutamátového receptoru? Receptor AMPA se podílí na synaptickém přenosu a je aktivován pouhým navázáním agonisty na vazebné místo. Naproti tomu receptor NMDA, jehož kanál je při přenosu podnětu s nízkou depolarizací blokován magnéziem, ke své aktivaci potřebuje jak navázání agonisty, tak silnou depolarizaci neuronu, která odstraní magnézium z kanálu. Blokáda magnéziem tak omezuje aktivaci receptoru NMDA i vápníkovou signalizaci pouze na specifické situace a chrání neuron před trvalým vstupem neurotoxického vápníku do buňky.Obecně se předpokládá, že při dlouhodobé potenciaci a dlouhodobé depresi přívodní podnět depolarizuje postsynaptický neuron do té míry, že magnézium je odstraněno z kanálu receptoru NMDA, který je tím aktivován. To vede ke vstupu vápníku do neuronů a dále k aktivaci nitrobuněčných systémů druhých poslů, které pak obměňují synaptický přenos. Jak je však možné, že aktivace receptoru NMDA během podněcování dlouhodobé potenciace a dlouhodobé deprese může různě zvýšovat koncentrace vápníku v buňce, který následně vyústí v dvousměrnou změnu síly synaptického přenosu? Odpověď zatím neznáme.
Možné zdroje vápníku
- Aktivace receptorů NMDA není jediný způsob jak zvýšit koncentraci vápníku v neuronu. Další cestou, kterou vápník do neuronu vstupuje, jsou napěťově závislé vápníkové kanály. Je popsáno několik typů těchto receptorů, které jsou stejně jako receptor NMDA aktivovány depolarizací postsynaptického neuronu. Zdá se však, že tyto vápníkové kanály hrají klíčovou úlohu pouze v synaptické plasticitě, která je na receptorech NMDA nezávislá.
- Dalším možným zdrojem vápníku jsou zásobárny uvnitř neuronu, z nichž je vápník uvolňován dvojím způsobem. Jednak zvýšení koncetrace vápníku v neuronu vyvolá jeho uvolnění v zásobárnách přes speciální receptory. Ukázalo se, že blokátor těchto receptorů brání indukci dlouhodobého zesílení synaptického přenosu. Účinek na dlouhodobé zeslabení přenosu dosud nebyl zkoumán. Pokud by však aktivace receptoru NMDA během zesílení a zeslabení přenosu podnětu vedla ke stejnému nárůstu vápníku v buňce, lze si těžko představit, že tento mechanizmus by umožnil uvolnění vápníku ze zásobáren v neuronu při potenciaci a znemožnil při depresi. Druhý způsob uvolňování vápníku ze zásobáren neuronu je přes aktivaci některých podtypů glutamátového receptoru. Navázání agonisty vede k aktivaci enzymu, který štěpí jeden z membránových lipidů a z nitrobuněčných zásobáren se uvolňuje vápník. Výzkum úlohy receptoru v zesílení a zeslabení synaptického přenosu však doposud nepřinesl žádná data svědčící pro rozdílnou aktivaci receptoru.
- Není vyloučeno, že zmíněné mechanizmy přispívají k zvýšení koncentrace nitrobuněčného vápníku při indukci synaptické plasticity. Žádný z nich však neposkytuje vysvětlení pro skutečnost, že nárůst vápníkových iontů je vyšší u dlouhodobé potenciace a nižší u dlouhodobé deprese. Odpověď na tuto otázku může být v samotném receptoru NMDA, který se – obdobně jako řada dalších receptorů – skládá z podjednotek. Dnes jich známe celkem pět: NR1 a čtyři NR2 (A až D). Studie s radioaktivně označenými agonisty a antagonisty ukázaly, že v mozku existují čtyři typy receptorů NMDA. Jednotlivé typy NMDA by se mohly odlišně podílet na indukci dlouhodobé potenciace a dlouhodobé deprese. U dlouhodobé deprese nízkofrekvenční podnět, který jenom částečně depolarizuje postsynaptickou membránu, může být dostatečný pro aktivaci receptorů obsahujících atypické podjednotky NR2C a NR2D. Při dlouhodobé vysokofrekvenční stimulaci depolarizuje membránu silněji a uvolní blok i z receptorů obsahujících NR2A a NR2B, které pak přispějí ke vstupu většího množství vápníkových iontů do buněk. Receptory obsahující podjednotky NR2C a NR2D jsou velmi pravděpodobně rovněž otevřeny při vysokofrekvenční stimulaci, ale pro LTP indukci nejsou samy o sobě dostačující.
Molekulární mechanizmus zesílení a zeslabení synaptického přenosu
Další klíčovou otázkou synaptické plasticity je molekulární mechanizmus dlouhodobé potenciace a dlouhodobé deprese (tab.). Aktivace receptorů během indukční fáze synaptické plasticity vede k aktivaci širokého spektra transdukčních (přenosových) molekul a druhých poslů. U dlouhodobé potenciace vedle již zmíněného nárůstu v nitrobuněčné koncentraci vápníku je to například nárůst kyseliny arachidonové a oxidu dusnatého, a aktivace fosforylujících enzymů, jako jsou kalcium/kalmodulinkináza typu II (CaMKII), cAMPkináza, tyrozinkináza a proteinkináza C (PKC). Antagonisté receptorů a inhibitory výše zmíněných molekul brání vzniku dlouhodobé potenciace. Během indukční fáze jsou také spuštěny procesy vedoucí k syntéze řady proteinů. Indukční fáze dlouhodobé deprese vyžaduje (stejně jako dlouhodobá potenciace) aktivaci receptorů NMDA, zvýšení oxidu dusnatého (NO) a aktivaci CaMkinázy II. Navíc však vyžaduje aktivaci několika defosforylujících enzymů, proteinfosfatáz. Indukce jak dlouhodobého zesílení přenosu, tak jeho zeslabení, je tedy velmi složitá. Naproti tomu druhá fáze synaptické plasticity – udržovací fáze – se zdá být mnohem jednodušší. V roce 1989 formuloval John Lisman hypotézu, ve které spojuje dlouhodobou potenciaci se zvýšenou úrovní fosforylace, zatímco dlouhodobou depresi se snížením fosforylace. Jeho hypotéza byla později experimentálně potvrzena v řadě laboratoří.
a druhých poslů. U dlouhodobé potenciace vedle již zmíněného nárůstu v nitrobuněčné koncentraci vápníku je to například nárůst kyseliny arachidonové a oxidu dusnatého, a aktivace fosforylujících enzymů, jako jsou kalcium/kalmodulinkináza typu II (CaMKII), cAMPkináza, tyrozinkináza a proteinkináza C (PKC). Antagonisté receptorů a inhibitory výše zmíněných molekul brání vzniku dlouhodobé potenciace. Během indukční fáze jsou také spuštěny procesy vedoucí k syntéze řady proteinů. Indukční fáze dlouhodobé deprese vyžaduje (stejně jako dlouhodobá potenciace) aktivaci receptorů NMDA, zvýšení oxidu dusnatého (NO) a aktivaci CaMkinázy II. Navíc však vyžaduje aktivaci několika defosforylujících enzymů, proteinfosfatáz. Indukce jak dlouhodobého zesílení přenosu, tak jeho zeslabení, je tedy velmi složitá. Naproti tomu druhá fáze synaptické plasticity – udržovací fáze – se zdá být mnohem jednodušší. V roce 1989 formuloval John Lisman hypotézu, ve které spojuje dlouhodobou potenciaci se zvýšenou úrovní fosforylace, zatímco dlouhodobou depresi se snížením fosforylace. Jeho hypotéza byla později experimentálně potvrzena v řadě laboratoří.
Vynořuje se však mnoho dalších otázek: Co je substrátem pro aktivované kinázy a fosfatázy? Jsou to receptory, či strukturální proteiny? Dochází k těmto změnám v presynaptickém, nebo v postsynaptickém neuronu? Podílí se na expresi synaptické plasticity nějaký zpětný posel? Vyřešení těchto otázek nám v budoucnu možná přinese odpověď na otázku nejdůležitější: Je synaptická plasticita opravdu mechanizmem, který mozek používá k tvorbě a ukládání paměti?
Literatura
Bliss T. V. P., Collingridge G. L.: A synaptic model of memory: long-term potentiation in the hippocampus, Nature 361, 31–39, 1993Dudek S. M., Bear M. F.: Homosynaptic long-term depression in area CA1 of hippocampus and effects of N-methyl-D-aspartate receptor blockade, Proc. Natl. Acad. Sci. USA 89, 4363–4367, 1992
Hrabetova S., Sacktor T. C.: Long term potentiation and long-term depression are induced through pharmacologically distinct NMDA receptors, Neurosci. Lett. 226, 107–110, 1997
Schwartz J. H.: Cognitive kinases, Proc. Natl. Acad. Sci. USA 90, 8310–8313, 1993
Stevens C. F.: Spatial lerning and memory: the beginning of a dream, Cell 81,1147–1148, 1996
Obrázky




axony – (ř. axon – osa) výběžky nervové buňky obsahující axoplazmu, zde axony granulárních buněk ve spoji synaptického okruhu, zvané šplhavá vlákna
agonista – (ř. agonistes – zápasník) zde ve významu stejně působící činitel
antagonista – (ř. antagonistes – odpůrce) opačně působící činitel
depolarizace nervového vlákna – změna membránového potenciálu vlákna po podráždění
indukce nervového vlákna – (lat. inducere – uvádět) zde část procesu vedoucího k diferenciaci
inhibitor – (lat. inhibere – zadržovat, zastavovat) tlumič, látka zpomalující reakci
ANATOMIE HIPOKAMPU
Hipokampová formace se skládá z gyrus dentatus, oblasti CA3, oblasti CA1 a subicula (obr. obrázek). Klasický třísynaptický okruh tvoří dopředná dráha, která začíná v čichové oblasti mozkové kůry a propojuje hlavní části hipokampové formace. První spoj třísynaptického okruhu začíná v mozkové kůře a končí synapsemi na granulárních buňkách gyrus dentatus. Druhý spoj je tvořen axony granulárních buněk (tzv. šplhavými vlákny), které mají zakončení na pyramidových neuronech v oblasti CA3. Axony CA3 pyramidových neuronů (Schafferovy odbočky) pak představují třetí spoj v okruhu a končí synapsemi na pyramidových neuronech oblasti CA1. Tento klasický model hipokampového okruhu byl doplněn nálezem vláken vedoucích z oblasti CA1 do subikula (od subicere – podkládat) a kůry, která de facto uzavírají smyčku okruhu. Synaptická plasticita je však v různých formách přítomna na všech synapsích základního třísynaptického okruhu.
HLAVNÍ RYSY SYNAPTICKÉ PLASTICITY
- Dlouhodobé zesílení synaptického přenosu (LTP) je rychle vyvolatelné, přitom výsledná změna synaptického přenosu trvá dlouho a vztahuje se pouze na synapse, které byly stimulovány.
- Zesílení synaptického přenosu je asociativní. To znamená, že společná aplikace silného a slabého podnětu postsynaptického neuronu přemění slabý podnět, který by samostatně žádnou trvalejší změnu synaptického přenosu nevyvolal, v podnět vzbuzující dlouhotrvající zesílení.
- Výskyt zesíleného synaptického přenosu je vázán na mozkové struktury zapojené do paměťových procesů. Kromě hipokampu je to například striatum podílející se na učení nových motorických dovedností nebo mozková kůra, která je zapojena do mnoha procesů souvisejících s tvorbou a ukládáním paměti.
Neuronové synapse
 Synapse jsou místa, na kterých dochází k přenosu elektrických signálů mezi nervovými buňkami. Běžná představa o tvaru synapse je ovlivněna zjednodušenými učebnicovými schématy (viz také naše schéma obrázek). Ve skutečnosti jsou však synapse a struktury s nimi spojené tvarově mnohem rozmanitější. Lze je vizualizovat a měřit díky rekonstrukcím vytvořeným ze sériových snímků z elektronového a konfokálního mikroskopu. Neocenitelnou pomoc při tom poskytují počítačové grafické systémy. Počítačové rekonstrukce jsou dnes jedinou metodou, která dovoluje takové zobrazení získat.
Synapse jsou místa, na kterých dochází k přenosu elektrických signálů mezi nervovými buňkami. Běžná představa o tvaru synapse je ovlivněna zjednodušenými učebnicovými schématy (viz také naše schéma obrázek). Ve skutečnosti jsou však synapse a struktury s nimi spojené tvarově mnohem rozmanitější. Lze je vizualizovat a měřit díky rekonstrukcím vytvořeným ze sériových snímků z elektronového a konfokálního mikroskopu. Neocenitelnou pomoc při tom poskytují počítačové grafické systémy. Počítačové rekonstrukce jsou dnes jedinou metodou, která dovoluje takové zobrazení získat.
Josef Špaček
Ke stažení
 Článek ve formátu PDF [167,53 kB]
Článek ve formátu PDF [167,53 kB]
O autorech
Sabina Hrabětová
Richard Rokyta
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
