









Od buněk k molekulám
 Imunitní systém brání organizmus proti cizorodým látkám a mikroorganizmům. Zneškodňuje abnormální vlastní buňky, které by pro organizmus mohly být nebezpečné. Celý obranný systém musí být schopen rozeznávat „normální“ složky organizmu od složek abnormálních a cizorodých. Tomu slouží nespecifické mechanizmy, jimž jsme se věnovali minule (Vesmír 73, 615, 1994/11), a mechamizmy antigenně specifické.
Imunitní systém brání organizmus proti cizorodým látkám a mikroorganizmům. Zneškodňuje abnormální vlastní buňky, které by pro organizmus mohly být nebezpečné. Celý obranný systém musí být schopen rozeznávat „normální“ složky organizmu od složek abnormálních a cizorodých. Tomu slouží nespecifické mechanizmy, jimž jsme se věnovali minule (Vesmír 73, 615, 1994/11), a mechamizmy antigenně specifické.
{kind=link}
Antigenně specifické mechanizmy
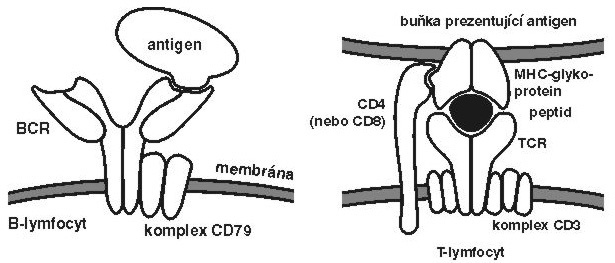
jsou vývojově dokonalejší (Vesmír 70, 625, 1991/11) a založeny na tom, že dva hlavní druhy lymfocytů, B a T, disponují antigenně specifickými receptory. Každý lymfocyt B nebo T (přesněji každý klon – skupina buněk pocházející z jediné původní buňky) nese na svém povrchu pouze jeden typ těchto receptorů, který se od receptorů na ostatních buňkách odlišuje detaily struktury vazebného místa pro antigen.Antigenně specifickými receptory B-lymfocytů jsou povrchové imunoglobuliny (sIg), kterým se také někdy říká B-receptory (BCR). V případě T-lymfocytů se obdobné molekuly nazývají T-receptory (TCR). Tyto názvy samozřejmě nesmějí svádět k představě, že BCR nebo TCR jsou jediné povrchové receptorové molekuly těchto buněk – na povrchu všech druhů buněk jsou stovky až miliony kopií desítek, či spíše stovek různých druhů jiných receptorových molekul (pro cytokiny, adhezivní molekuly a další). Imunitní systém disponuje obrovským repertoárem (nejméně 100 milionů) odlišných klonů lymfocytů B a T, jinak řečeno bezpočtem struktur a specifit vazebných míst jejich antigenně specifických receptorů.
Tak velká strukturní rozmanitost je dána hlavně (ale ne výlučně) unikátním genetickým mechanizmem vcelku náhodného kombinování malých genových úseků kódujících variabilní části polypeptidových řetězců těchto receptorů během vývoje a zrání lymfocytů (obrázek). etailnější popis tohoto neobyčejně zajímavého procesu by vyžadoval samostatný článek.
{kind=link}
Antigenně specifický imunitní systém je tedy hlavně systémem anticipačním. V novorozeném organizmu, který se ještě nesetkal s žádným cizorodým antigenem, je předem připraven „pro všechny případy“ tak velký sortiment specifit, že se v něm najdou receptory schopné vázat téměř jakoukoli chemickou strukturu. Situace připomíná velký konfekční obchod, v němž jakýkoli zákazník najde šaty, které mu dobře padnou. Později uvidíme, že imunologický konfekční obchod, alespoň jeho B-lymfocytární část, zaměstnává také krejčího, který dodatečně upravuje vybrané obleky tak, aby padly ještě lépe.
 Existuje základní rozdíl mezi antigenně specifickými receptory lymfocytů B a T: zatímco BCR rozpoznávají antigeny jako takové, TCR rozlišují komplexy malých peptidových fragmentů antigenů a MHC glykoproteinů (major histocompatibility complex; známé též jako histokompatibilitní antigeny nebo transplantační antigeny, Vesmír 69, 274, 1990/5; obrázek). Součástí TCR jsou ještě další koreceptorové molekuly CD4 a CD8 (podle typu T-buněk), které posilují vazbu mezi TCR a rozeznávaným komplexem MHC-glykoproteinu s peptidem.
Existuje základní rozdíl mezi antigenně specifickými receptory lymfocytů B a T: zatímco BCR rozpoznávají antigeny jako takové, TCR rozlišují komplexy malých peptidových fragmentů antigenů a MHC glykoproteinů (major histocompatibility complex; známé též jako histokompatibilitní antigeny nebo transplantační antigeny, Vesmír 69, 274, 1990/5; obrázek). Součástí TCR jsou ještě další koreceptorové molekuly CD4 a CD8 (podle typu T-buněk), které posilují vazbu mezi TCR a rozeznávaným komplexem MHC-glykoproteinu s peptidem.
{kind=link}
Tento rozdíl mezi BCR a TCR souvisí se základní strategií B- a T-lymfocytů: Zatímco B-lymfocyty se specializují na produkci protilátek, které se vážou na cizorodé látky (antigeny) rozpuštěné v tělních tekutinách nebo deponované na povrchu mikroorganizmů, T-buňky se zaměřují výhradně na rozeznávání antigenů nebo jejich fragmentů na povrchu vlastních buněk infikovaných viry nebo nitrobuněčnými parazity.
 Antigenně specifické mechanizmy jsou tedy založeny na tom, že lymfocyty, jež něco rozeznaly, se množí (vytvoří klon mnoha buněk s identickou specifitou) a diferencují se v efektorové buňky, které v optimálním případě zneškodní rozeznaný antigen či mikroorganizmus. V takovém systému musí ovšem existovat dokonalé mechanizmy odstraňování nebo alespoň blokování funkce lymfocytů, které by rozeznávaly a napadaly struktury a buňky organizmu vlastní.
Antigenně specifické mechanizmy jsou tedy založeny na tom, že lymfocyty, jež něco rozeznaly, se množí (vytvoří klon mnoha buněk s identickou specifitou) a diferencují se v efektorové buňky, které v optimálním případě zneškodní rozeznaný antigen či mikroorganizmus. V takovém systému musí ovšem existovat dokonalé mechanizmy odstraňování nebo alespoň blokování funkce lymfocytů, které by rozeznávaly a napadaly struktury a buňky organizmu vlastní.
Antigenně specifický imunitní systém lze rozdělit na část zodpovědnou za reakce:
- humorálně (protilátkově) zprostředkované a
- buněčně zprostředkované.
 Tyto dvě větve imunitního systému jsou si v mnohém podobné, ale v mnohém se liší.
Tyto dvě větve imunitního systému jsou si v mnohém podobné, ale v mnohém se liší.
{kind=link}
Humorální (protilátková) imunitní odpověď
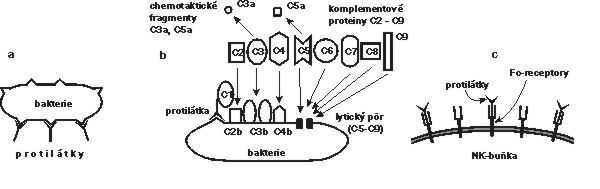
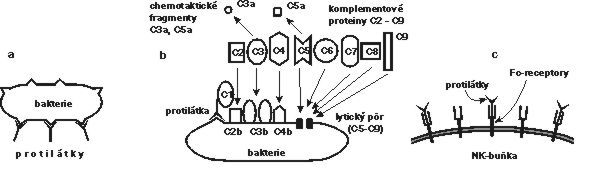
Strategie této části imunitního systému spočívá ve vytvoření velkého množství rozpustných protilátek – imunoglobulinů, které se specificky vážou na cizorodé molekuly a na struktury invazních mikroorganizmů. Některé z těchto protilátek mohou blokovat („neutralizovat“) škodlivé působení cizorodých látek nebo mikroorganizmů již samotným navázáním, a to pokud se např. vážou na aktivní místo bakteriálního toxinu nebo na místo, jímž se mikroorganizmus uchycuje na povrchu infikované buňky. Většinou však samotná vazba protilátky ještě cizorodý antigen nebo mikroorganizmus nezneškodní a je potřeba dalších efektorových mechanizmů (obrázek).Jedním z nich je opsonizační efekt – částice „označená“ protilátkami se naváže na Fc-receptory fagocytujících buněk a je jimi pozřena. Druhým hlavním efektorovým mechanizmem je již zmíněná aktivace komplementové kaskády (Vesmír 73, 615, 1994/11) prostřednictvím molekul protilátek navázaných na antigenní částici.
 Další efektorový mechanizmus, jehož skutečný význam in vivo není docela jasný, představuje buněčná cytotoxicita závislá na protilátkách (ADCC; antibody-dependent cell cytotoxicity). Na abnormální buňky (nádorové, infikované viry) obalené protilátkami třídy IgG se prostřednictvím Fc-receptorů mohou navázat NK-buňky (které, jak ještě uvidíme, mají i jiné funkce) a mohou je zahubit kontaktními cytotoxickými mechanizmy.
Další efektorový mechanizmus, jehož skutečný význam in vivo není docela jasný, představuje buněčná cytotoxicita závislá na protilátkách (ADCC; antibody-dependent cell cytotoxicity). Na abnormální buňky (nádorové, infikované viry) obalené protilátkami třídy IgG se prostřednictvím Fc-receptorů mohou navázat NK-buňky (které, jak ještě uvidíme, mají i jiné funkce) a mohou je zahubit kontaktními cytotoxickými mechanizmy.
{kind=link}
Velmi podobně působí protilátky třídy IgE – ty se mohou pevně navázat na vysokoafinní IgE-receptory na povrchu bazofilů a tzv. žírných buněk. Tímto způsobem se žírné buňky zprostředkovaně vyzbrojí antigenně specifickými receptory. Setkají-li se pak s částicí nesoucí příslušný antigen, navážou se na ni a IgE-receptory předají o této události do nitra buňky signál. To vede k degranulaci – vylití obsahu cytoplazmatických váčků plných různých látek vyvolávajících místní zánětlivou reakci (obrázek). Tento způsob obrany je zvláště účinný proti některým mnohobuněčným parazitům. Imunitní systém bohužel často obdobně reaguje na mnohé neškodné částice, jako např. pylová zrnka. Výsledkem tohoto zbytečného boje jsou známé alergické reakce.
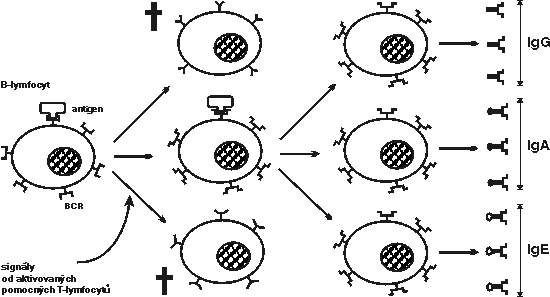
Tvorba protilátek specificky reagujících s určitým antigenem
Setká-li se B-lymfocyt poprvé ve svém životě s antigenem (Vesmír 71, 388, 1992/7), jehož nějaká struktura (epitop) vytváří dostatečně stabilní komplex s vazebným místem jeho povrchového imunoglobulinu, obdrží tak první, nezbytně nutný, ale většinou ještě nepostačující aktivační signál. Pokud v krátké době neobdrží ještě další pomocné signály, má tento první signál obdržený prostřednictvím povrchového imunoglobulinu výrazně negativní efekt: B-lymfocyt se na dlouhou dobu dostane do stavu anergie – neodpovídavosti – a v některých případech dokonce hyne (Vesmír 73, 550, 1994/10, Vesmír 73, 552, 1994/10). To je důležitý způsob zabraňující aktivaci potenciálně autoreaktivních B-lymfocytů a produkci vlastních protilátek. Pokud se B-lymfocyt, který dostal první signál rozeznáním antigenu, brzy setká s aktivovaným pomocným T-lymfocytem, dostane od něj pomocné signály nezbytné k pomnožení klonu a konečné přeměně v plazmatické buňky sekretující velká množství protilátek.
Pokud se B-lymfocyt, který dostal první signál rozeznáním antigenu, brzy setká s aktivovaným pomocným T-lymfocytem, dostane od něj pomocné signály nezbytné k pomnožení klonu a konečné přeměně v plazmatické buňky sekretující velká množství protilátek.
Co jsou ony pomocné signály a jak je B-lymfocyty přijímají? Jeden z nich obdrží B-lymfocyt, když dojde k vazbě mezi jeho povrchovým receptorem CD40 a membránovým glykoproteinem z povrchu aktivovaného pomocného T-lymfocytu, kterému se říká buď gp39, nebo CD40L (ligand CD40). Další pomocné signály dostává B-lymfocyt prostřednictvím specifických receptorů pro cytokiny IL-4 a IL-5, jež produkují také aktivované pomocné T-lymfocyty (obrázek). 1)
Ve stručném přehledu není dostatek prostoru na popis mechanizmů zajišťujících přenos signálů všemi uvedenými typy receptorů. Stačí jen říci, že základní úlohu má rychlá aktivace několika druhů kináz, tj. enzymů přenášejících fosfátovou skupinu z ATP na některé nitrobuněčné proteiny (jimiž jsou často další kinázy) a membránové lipidy. Tyto kinázy jsou nekovalentně asociovány s nitrobuněčnými částmi receptorových molekul a aktivují se konformačními změnami, resp. agregací receptorů, způsobenými navázáním ligandů.
 Dostane-li lymfocyt B všechny náležité signály, začne se dělit a vzniklý klon začne procházet mimořádně důležitým obdobím – fází, kdy dosud nejasným mechanizmem dojde k velmi intenzivním mutacím ve variabilních částech jejich genů kódujících imunoglobulinové řetězce a k produkci mutantních forem povrchových imunoglobulinů (obrázek). Jednotlivé mutantní formy se liší svou afinitou (silou vazby) k antigenu a důležité je, že přežívají pouze ty buňky, které jsou schopny vázat antigen co nejpevněji. Ostatní mutantní B-lymfocyty, které jej vážou slabě nebo vůbec ne, během tohoto procesu, zvaného afinitní maturace, hynou. Výsledkem afinitní maturace je vznik optimalizované varianty původní buňky, schopné produkovat velmi kvalitní vysokoafinní protilátky. Proces afinitní maturace je tedy oním zakázkovým krejčím, který přešívá konfekční oblek (protilátku) tak, aby zákazníkovi (antigenu) padl co nejdokonaleji.
Dostane-li lymfocyt B všechny náležité signály, začne se dělit a vzniklý klon začne procházet mimořádně důležitým obdobím – fází, kdy dosud nejasným mechanizmem dojde k velmi intenzivním mutacím ve variabilních částech jejich genů kódujících imunoglobulinové řetězce a k produkci mutantních forem povrchových imunoglobulinů (obrázek). Jednotlivé mutantní formy se liší svou afinitou (silou vazby) k antigenu a důležité je, že přežívají pouze ty buňky, které jsou schopny vázat antigen co nejpevněji. Ostatní mutantní B-lymfocyty, které jej vážou slabě nebo vůbec ne, během tohoto procesu, zvaného afinitní maturace, hynou. Výsledkem afinitní maturace je vznik optimalizované varianty původní buňky, schopné produkovat velmi kvalitní vysokoafinní protilátky. Proces afinitní maturace je tedy oním zakázkovým krejčím, který přešívá konfekční oblek (protilátku) tak, aby zákazníkovi (antigenu) padl co nejdokonaleji.
{kind=link}
Téměř zároveň často dochází k dalšímu přeskupení části imunoglobulinového genu a tím ke změně struktury imunoglobulinu – izotypovému přesmyku, při kterém se mění izotyp (třída resp. podtřída) těžkého řetězce imunoglobulinu z původního IgM nebo IgD na některý jiný (IgA, IgE nebo některou podtřídu IgG). Tato změna nijak neovlivňuje vazebnou specifitu protilátky, protože se odehrává na druhém konci molekuly, ale zásadním způsobem mění možnosti efektorových funkcí protilátek (schopnost vazby na Fc-receptory, opsonizace, aktivace komplementu). O tom, k jakému konkrétnímu izotypovému přesmyku dojde, rozhoduje pravděpodobně vzájemný poměr místních koncentrací několika kritických cytokinů (IL-4, IL-10, TGF-β, interferon-γ; Vesmír 71, 509, 1992/9).
Po afinitní maturaci a izotypovém přesmyku dokončí B-lymfocyty svůj vývoj a přemění se v plazmatické buňky – v podstatě malé továrny masově produkující vysoce specifické rozpustné protilátky, strukturně téměř stejné jako optimalizované povrchové imunoglobuliny. Část B-lymfocytů nedokončí vývoj až do stadia plazmatických buněk, ale stane se paměťovými B-lymfocyty. Dojde-li v budoucnosti k opakovanému setkání se stejným antigenem, aktivují se paměťové buňky již mnohem snáze. Jsou méně náročné na poskytnutí pomocných signálů a nemusejí už procházet fází afinitní maturace, proto dávají rychle vzniknout klonu plazmatických buněk. To je podstata známého průběhu sekundární protilátkové odpovědi a blahodárného účinku očkování. Zdá se, že pro udržování paměťových B-lymfocytů má velký význam podivuhodná schopnost folikulárních dendritických buněk skladovat na svém povrchu velmi dlouhou dobu (snad i roky!) antigeny ve formě komplexů s protilátkami.
Kde se však vezmou klíčově důležité aktivované pomocné T-lymfocyty? Nejčastěji vznikají z prekurzorových T-buněk poté, kdy tyto buňky rozeznaly pomocí svých T-receptorů peptidové fragmenty antigenu navázané na MHC-glykoproteiny II. třídy na povrchu buněk prezentujících antigen (APC; antigen presenting cells, obrázek). Pomocné T-buňky a jejich prekurzory nesou na svém povrchu, kromě mnoha jiných druhů molekul, také pro ně charakteristický glykoprotein CD4, který funguje jako koreceptor MHC-molekul II. třídy a posiluje vazbu T-receptoru. Účinnými APC jsou makrofágy (Vesmír 72, 252, 1993/5), a hlavně různé druhy dendritických buněk. Pokud tyto buňky pohltí antigenní částice nebo mikroorganizmus, rozštěpí jeho proteiny v lyzozomech na peptidové fragmenty, z nichž některé vytvoří komplexy s molekulami MHC-glykoproteinů II. třídy. Tyto komplexy pak buňky prezentující antigen „vystrčí“ na svůj povrch a předkládají je k rozeznání prekurzorům pomocných T-lymfocytů.
{kind=link}
 Mezi obrovským množstvím těchto buněk se tu a tam najde taková, jejíž T-receptory mohou vázat tento jeden určitý komplex MHC-glykoproteinu s antigenním peptidem a tak obdržet první nezbytný (ale opět nikoli postačující) aktivační signál. Stejně jako je tomu u B-lymfocytů, musí takový T-lymfocyt dostat ještě druhý signál (přesněji druhé signály), a to ve formě vazebných interakcí několika párů povrchových adhezivně-signalizačních molekul T-buňky a buňky prezentující antigen. Zdá se, že nejdůležitějším receptorem druhého signálu je pro T-lymfocyty povrchový protein CD28, který váže ligand B7 (CD80) přítomný na povrchu buněk prezentujících antigen. Kromě těchto adhezivních pomocných signalizačních molekul jsou k plné aktivaci pomocných T-buněk důležité také rozpustné cytokiny produkované buňkami prezentujícími antigen, zvláště interleukin 1 (IL-1).
Mezi obrovským množstvím těchto buněk se tu a tam najde taková, jejíž T-receptory mohou vázat tento jeden určitý komplex MHC-glykoproteinu s antigenním peptidem a tak obdržet první nezbytný (ale opět nikoli postačující) aktivační signál. Stejně jako je tomu u B-lymfocytů, musí takový T-lymfocyt dostat ještě druhý signál (přesněji druhé signály), a to ve formě vazebných interakcí několika párů povrchových adhezivně-signalizačních molekul T-buňky a buňky prezentující antigen. Zdá se, že nejdůležitějším receptorem druhého signálu je pro T-lymfocyty povrchový protein CD28, který váže ligand B7 (CD80) přítomný na povrchu buněk prezentujících antigen. Kromě těchto adhezivních pomocných signalizačních molekul jsou k plné aktivaci pomocných T-buněk důležité také rozpustné cytokiny produkované buňkami prezentujícími antigen, zvláště interleukin 1 (IL-1).
Pro nastartování humorální části imunitní odpovědi je potřeba souhry alespoň tří druhů buněk: B-lymfocytů, pomocných T-lymfocytů a buněk prezentujících antigen. Zdá se ovšem velmi nepravděpodobné, že by se tyto tři typy buněk sešly na jednom místě. Tato nepravděpodobná setkání jsou však velmi usnadněna tím, že se popisované děje odehrávají v anatomicky optimálně organizovaném prostředí lymfatických uzlin a jiných sekundárních lymfoidních orgánů. Zde se jednak na povrchu folikulárních dendritických buněk (FDC) koncentrují antigeny (ve formě komplexů s protilátkami a komplementovými fragmenty), jednak do nich přicházejí makrofágy a dendritické buňky se svým natráveným úlovkem a předkládají ho na svém rozsáhlém členitém povrchu T-lymfocytům, a konečně jsou zde nahloučeny ve velkém množství lymfocyty B i T. V zárodečných centrech zde probíhají děje afinitní maturace a kvalita mutantních forem povrchových imunoglobulinů je testována kontaktem s antigenem skladovaným na povrchu folikulárních dendritických buněk.
Někdy je celý systém jednodušší a vystačí s interakcí pouze dvou typů buněk: B-lymfocytů a pomocných T-lymfocytů. To se může, alespoň teoreticky, stát tehdy, když B-lymfocyt rozezná nějaký malý antigen, např. cizorodý protein. Ten pak může pohltit, rozštěpit a některé štěpy vystavit na svém povrchu v komplexu s MHC-glykoproteiny II. třídy, kde je mohou rozeznat pomocné T-lymfocyty. V tomto případě B-lymfocyt funguje současně jako buňka prezentující antigen a obě buňky (B a T) se vzájemně stimulují. Tento tzv. minimální model spolupráce se ovšem nemůže uplatnit v případě velkých antigenních částic, jako mikrobů, které B-lymfocyt pohltit a zpracovat nedokáže.
Ještě jednou připomeňme, jak je bráněno vzniku autoprotilátek (tj. protilátek reagujících se strukturami organizmu vlastními). V prvé řadě, v časném vývojovém stadiu, kdy nezralé B-lymfocyty mají na svém povrchu antigenně specifické receptory třídy IgM, vede setkání se strukturou schopnou vazby na receptor buď k odumření, nebo k dlouhotrvajícímu útlumu B-lymfocytu. Takto se eliminuje nebo dlouhodobě zneškodňuje velká většina potenciálně autoreaktivních buněk, které se setkají s autoantigenem (tj. např. sérovými proteiny nebo povrchovými molekulami různých buněk). Pokud k setkání s autoantigenem dojde až ve stadiu zralého B-lymfocytu, který většinou nese antigenně specifické receptory třídy IgD, je k plné aktivaci nutná spolupráce pomocných T-lymfocytů. Pomocné T-buňky, které by rozeznávaly fragmenty autoantigenu, však byly eliminovány během vývoje v tymu (Vesmír 69, 391, 1990/7). Nedostane-li tedy potenciálně autoreaktivní zralá B-buňka pomoc od T-lymfocytů, dostane se i ona do stavu funkčního útlumu (anergie) a nemůže začít produkovat autoprotilátky.
Poznámky
antigen – látka (obvykle cizorodá) schopná za vhodných okolností vyvolat v organizmu imunitní odpověď
autoantigen – látka organizmu vlastní, která však za určitých okolností může vyvolat imunitní odpověď; je příčinou autoimunitních onemocnění
autoreaktivní buňky – buňky imunitního systému, které rozeznávají autoantigeny a snaží se je ničit
imunokomplex – složitá funkční struktura z molekul účastnících se obranné reakce, především antigenu a protilátky
klonování – metodické postupy umožňující získání homogenních skupin buněk – klonů, nebo replikujících se genetických elementů pocházejících z jedné původní rodičovské buňky, resp. molekuly
konformační změna – změna tvaru molekuly
koreceptor – molekula pomáhající funkci receptorové molekuly
ligand – molekula, která se váže na receptor nebo jinou vazebnou molekulu
prekurzorové buňky – buňky, které jsou ve vývoji předchůdci jiných buněk; tymocyty jsou prekurzory T-lymfocytů, B-lymfocyty jsou prekurzory plazmatických buněk, monocyty jsou prekurzory makrofágů
protilátky (imunoglobuliny) – proteiny (bílkoviny) charakteristické struktury produkované B-lymfocyty (či spíše tzv. plazmatickými buňkami, které vznikají z B-lymfocytů); skládají se z tzv. lehkých (L) a těžkých (H) řetězců; vážou antigeny a tím je alespoň v některých případech zneškodňují nebo pomáhají jejich zneškodnění; principy vzniku protilátek a jejich strukturní a vazebné rozmanitosti jsou popsány v textu; existuje několik strukturně poněkud odlišných imunoglobulinů (tzv. třídy nebo izotypy IgM, IgD, IgG, IgA, IgE)
žírné buňky – zvláštní druh buněk roztroušených ve tkáních (např. ve sliznicích) vyznačující se přítomností velkého množství receptorů pro imunoglobuliny třídy IgE; tyto buňky jsou odpovědné za projevy alergií; jejich pozitivní úlohou je obrana proti parazitům
O autorovi
Václav Hořejší

Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
