










Morfologie milování
[...] Jsou to dva mořští koníčkové, jak kopulujou, a zeptám se vás, jestli na tom je něco zvláštního. Nevidíte nic, prima. Tadyhle ten takovej čudlík malinkej co vidíte, co vykukuje z toho jednoho zviřátka do druhýho, jak to vypadá jako nějakej kopulační intromitentní orgán, tak to skutečně kopulační intromitentní orgán je, jenomže je to orgán nikoli samčí, ale samičí. [...] (úryvek z přednášky)
Shrnutí předchozí části
Většina skupin živočichů s vnitřním oplodněním má složité a druhově specifické samčí genitálie (popřípadě genitálie sekundární, objímadla či spermatofory), zatímco samičí genitálie bývají jednoduché a uniformní, a příslušné druhy se často v jiných tělesných znacích neliší. Probrali jsme různé neobvyklosti a došli k závěru, že základní situaci lze vysvětlit v kontextu pohlavního výběru s přihlédnutím k odlišnosti samčích a samičích zájmů. Shrnuli jsme hypotézy vysvětlující samčí rozmanitost a samičí jednotvárnost (viz box 2 ), zamítli ty, které jsou založeny na představě „samičího zámku“ a „samčího klíče“ nebo vedlejšího pleiotropického efektu, a podpořili ty, které vycházejí z teorie samičí volby a zdůrazňují význam libida.Promiskuita samice
Základním předpokladem teorie samičí volby založené na selekci samců dle kvality jejich genitálií je jistý stupeň promiskuity samice. Má-li si totiž samice vybírat samce podle kvality stimulace, kterou jí poskytuje při kopulaci, musí „vyzkoušet“ alespoň několik samců. To je vlastně také jeden ze základních problémů celé teorie, neboť se ví jen málo o tom, kolikrát samice různých druhů v přírodě kopulují. Dobré znalosti máme u modelově studovaných druhů obratlovců, ale o milionech druhů bezobratlých nevíme téměř nic. Sledování promiskuity je navíc obtížný problém. V zajetí nelze nikdy vytvořit přirozené prostředí a v přírodě je téměř nemožné sledovat nějakého jedince hmyzu nebo třeba ploštěnky po celou dobu jejich aktivního sexuálního života. Podle záznamů u těch skupin, kde samec zanechává jasnou stopu po kopulaci (jizvy po traumatické inseminaci, ulomený konec makadla některých pavouků, který zůstává v samičí semenné schránce nebo zbytky samčího spermatoforu v těle samic některých motýlů), však docházíme k názoru, že většina druhů tendenci k samičí promiskuitě má.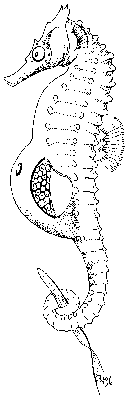 Přesto však můžeme – alespoň v některých případech – monogamii předpokládat. Často si monogamii samice snaží zajistit samci. U některých druhů vytvářejí kopulační uzávěry nazývané u hmyzu sfragmata a u obratlovců kopulační zátky. Myslelo se, že tato „zařízení“ bezpečně zajišťují samcům paternitu. Ale stejně jako ve středověku neplnily vždy pásy cudnosti svou roli, nefungují ani sfragmata či kopulační zátky stoprocentně. Bylo např. zjištěno, že v kopulačních orgánech samic jasoňů jsou spermie několika samců, i když ti vždy pečlivě umísťují sfragmata. O samicích hlodavců je známo, že si kopulační zátky samy aktivně odstraňují a mohou tak kopulovat s více samci.
Přesto však můžeme – alespoň v některých případech – monogamii předpokládat. Často si monogamii samice snaží zajistit samci. U některých druhů vytvářejí kopulační uzávěry nazývané u hmyzu sfragmata a u obratlovců kopulační zátky. Myslelo se, že tato „zařízení“ bezpečně zajišťují samcům paternitu. Ale stejně jako ve středověku neplnily vždy pásy cudnosti svou roli, nefungují ani sfragmata či kopulační zátky stoprocentně. Bylo např. zjištěno, že v kopulačních orgánech samic jasoňů jsou spermie několika samců, i když ti vždy pečlivě umísťují sfragmata. O samicích hlodavců je známo, že si kopulační zátky samy aktivně odstraňují a mohou tak kopulovat s více samci.
Opět se tu vlastně jen projevuje již zmiňovaný střet zájmů obou pohlaví – samec si chce zajistit paternitu, samice vybírá kvalitního otce pro své potomstvo.
Dále je tu vynucená monogamie, kdy určitému živočichovi okolnosti způsobu jeho života nic jiného vlastně ani nedovolí. Např. dospělci některých jepic žijí skutečně tak krátce, že prostě nemají čas seznámit se během svého reprodukčního života s více samci. U termitů je zase královna spolu s jediným samcem doslova obezděna v královské komůrce. Dokud samec nezemře, jiný se k ní nedostane. Termiti žijí opravdu v přísné monogamii. U obou jmenovaných skupin však nacházíme jednoduché a téměř uniformní genitálie. Intrasexuální selekce založená na genitáliích tu tedy ztrácí svůj význam a jednoduché genitálie u monogamních skupin její existenci nepřímo potvrzují.
Možnosti samice
O možnostech samičí volby při kopulaci se dovídáme až v poslední době. Samice může např. připustit samce hlouběji do svých genitálních cest, blíže k místům, kde jsou oocyty nebo vajíčka, která budou oplodňována. Žádná ze skupin s diverzifikovanými genitáliemi však neumísťuje spermie přímo na oocyty nebo na vajíčka. Podle stupně účinnosti samčích stimulů může samice přerušit kopulaci, zabránit inseminaci, dovolit vícenásobnou inseminaci; má prostě řadu možností, kterými ovlivňuje rozmnožovací úspěch samce již v kopulačně-inseminační fázi.Je důležité si uvědomit, že genitální stimulace může být pro samici jediným a výlučným důvodem k tomu, aby dala přednost jednomu určitému samci. Nepotřebují žádný estetický podnět, jak se domníval Darwin, který předpokládal, že u ptáků vyhledávají samice obzvláště krásně zbarvené samce, a tuto myšlenku zobecnil. Nepíšeme tu však o atraktivnosti obecně, jinak bychom o této situaci museli dlouze diskutovat. Právě ptáci se složitým předkopulačním rituálem a uniformními až téměř neexistujícími samčími genitáliemi jsou tím nejméně vhodným příkladem pro naše téma.
Jakým způsobem však může samice vyvolávat selekční tlak na genitálie samce? Představme si, že vznikne náhodná geneticky podložená struktura, která stimuluje samici poněkud více než obvykle. Samice dá přednost spermiím samce, který ji takto stimuloval, fertilizuje jimi vajíčka a gen pro samčí novinku se přenáší preferenčně do dalšího pokolení. Genitálie obou pohlaví totiž musí být v jisté korelaci, i když ne jen mechanicky, tak jak si mysleli tvůrci teorie zámku a klíče, ale spíše smyslově.
Ne každý sexuální kontakt, ne každá intromise vede nutně k inseminaci. Samice mnoha druhů (např. octomilky rodu Drosophila) provádějí před vlastní inseminační kopulací tzv. pseudokopulace, při nichž nikdy nedojde k inseminaci. Maximálně promiskuitní a přitom zároveň fertilizačně monogamní jsou samice mouchy domácí (Musca domestica) a některé druhy masařek a komárů. Kopulují totiž třeba až 200krát s různými samci celkem bez výběru, pseudokopulaci nepřerušují, ale žádnému samci nedovolí inseminaci. Až teprve samci, který zcela „vyhovuje jejím představám“, zhruba po 100 – 200 kopulacích, dovolí moucha inseminaci, která pravděpodobně vede i k fertilizaci, a právě tento samec se stává otcem potomstva. Zároveň však tento poslední samec do těla samice produkuje antiafrodiziakální látky, takže ostatní samci ztrácejí o samici zájem. Pozoruhodné je, že se vzrůstajícím stářím jsou samice stále méně vybíravé, a tedy paradoxně méně promiskuitní.
Samice mohou silně ovlivňovat i kvalitu kopulace a následné ovipozice. U mrchožravých srpic z rodu Panorpa (řád Mecoptera) je zvykem, že samec přináší samici před kopulací dárek – čerstvou mrtvolku hmyzu. Samec podle svých schopností mrtvolku buď najde, nebo ji ukradne pavoukovi ze sítě, anebo má ještě jednu možnost – místo mrtvolky může samičce naslintat proteinový balíček, který samice přijímá jako dárek rovnocenný oné mrtvolce. Pak jsou však ještě neschopní samci, kteří ani nenajdou, ani neukradnou mrtvolku a nedokáží ani naslintat náhradní dárek. Ti volí v živočišné říši téměř ojedinělou strategii a samici prostě znásilní. Znásilnění zde slouží jako pravidelná rozmnožovací strategie užívaná poměrně méně hodnotnými samci. Násilí ovšem sníží ovipozici samice asi o 50 %.
 Jak jsem se už zmínil, jsou u živočichů s diverzifikovanými genitáliemi spermie téměř vždy uloženy v těle daleko od vlastních vajíček. Samice má značné možnosti ovlivnit, jak blízko se ty které spermie při inseminaci dostanou. Může ovládat svou spermatéku (semennou schránku) a svalově nebo chemicky rozšiřovat či naopak zužovat své genitální cesty. Míra změny závisí do značné míry na vzrušení samice (bezpečně známo u savců). A naopak, kvalita kopulace ovlivňuje nejen vnitřní tvar samičích genitálií, ale i dozrávání vajíček, rychlost ovulace a ovipozice, množství výživy, která se dostane oocytu či zárodku, i rychlost cyklu, při němž se vnitřní orgány připravují na uhnízdění vajíčka u živočichů s placentou (někteří obratlovci) nebo pseudoplacentou (např. některý hmyz).
Jak jsem se už zmínil, jsou u živočichů s diverzifikovanými genitáliemi spermie téměř vždy uloženy v těle daleko od vlastních vajíček. Samice má značné možnosti ovlivnit, jak blízko se ty které spermie při inseminaci dostanou. Může ovládat svou spermatéku (semennou schránku) a svalově nebo chemicky rozšiřovat či naopak zužovat své genitální cesty. Míra změny závisí do značné míry na vzrušení samice (bezpečně známo u savců). A naopak, kvalita kopulace ovlivňuje nejen vnitřní tvar samičích genitálií, ale i dozrávání vajíček, rychlost ovulace a ovipozice, množství výživy, která se dostane oocytu či zárodku, i rychlost cyklu, při němž se vnitřní orgány připravují na uhnízdění vajíčka u živočichů s placentou (někteří obratlovci) nebo pseudoplacentou (např. některý hmyz).
Možnosti samce
Zatím vše vypadalo tak, že osudy samců jsou vlastně jen hříčkou v „rukou“ samic. Není tomu tak docela. Tak především samec může z těla samice odstranit spermie samce, který jej „předběhl“. Typickým příkladem jsou vážky, zejména šidélka. Mají jednoduché primární a složité sekundární genitálie většinou adaptované tak, že samec, který transportuje spermatofor do pohlavního otvoru samice, jako lžičkou vyškrábne spermatofor, který tam před ním umístil jiný samec.Dále se samec může pokusit o to, aby byl poslední, kdo se samicí kopuloval. Poslední (alespoň u hmyzu, u obratlovců je tomu jinak) má výhodu v tom, že jeho spermie se dostanou nejblíže místu, kudy bude procházet vajíčko, tj. k místu, kde bude oplozováno. Poslední samec také může produkovat spermicidní látky, které jsou individuálně odlišné a dokáží likvidovat spermie předchozích rivalů. Také zaplavení útrob samice velkým množstvím spermií zvýší jeho šanci oproti méně produktivnímu předchůdci.
Další možnosti poskytují samcům nejrůznější objímací orgány, kterými se přidržují těla samice. Donedávna se skutečně mělo za to, že slouží pouze mechanickému spojení během kopulace. Nově se však ukazuje, že navíc umožňují libidózní stimulaci samice. Samec tím může přimět samici (jestliže „se jí to líbí“), aby se nechala držet a samce ze sebe nesetřásla.
Pozoruhodná je strategie samců olihně (Loligo poelii, hlavonožci – Cephalopoda), kteří mohou do jisté míry ovlivnit samičí volbu spermií. Samice tohoto druhu kopuluje opakovaně s různými samci, kteří na různých místech v jejím těle ukládají spermatofory. V okamžiku, kdy samice začne snášet vajíčka, spermatofory vybuchují, a teprve tehdy dochází k oplodnění. Právě ten samec, který vystihne okamžik, kdy bude samice ovulovat a klást vajíčka, a v ten okamžik vystřelí do její plášťové dutiny explodující spermatofor, s největší pravděpodobností vajíčka oplodní.
Naprosto extrémní situací je traumatická inseminace, kdy se spermie nedostávají do samice obvyklou genitální cestou, ale samice je samčím intromitentním orgánem vždy zraněna. Např. u štěnic samec probodne membránu spojující články zadečku samice nebo kopuluje do uzavřeného orgánu samice umístěného daleko od vulvy. Kromě ploštic je traumatická inseminace známa i u řásníků (Strepsiptera), drápkovců (Onychophora), některých ploštěnek aj. U některých skupin mají samice dokonce vytvořeny nové, tzv. paragenitální cesty, kterými přivádějí spermie k vajíčkům. Traumatická inseminace je vždy spojena s tzv. „hypergamezí“, situací, kdy samec produkuje nadměrné množství spermií, z nichž podstatná část slouží výživě samice.
Traumatickou inseminaci začaly první druhy a první jedinci praktikovat asi proto, aby zkrátili cestu spermií složitými vnitřními genitáliemi samice (přesněji: bylo to pro samce selekčně výhodné). Spermie se tak vlastně „zkratkou“ dostanou rychle na správné místo a vyhnou se samičí volbě. „Na oplátku“ se však u samic opět vyvinula metoda, jak selektovat spermie. U samic druhů s traumatickou inseminací byly nalezeny spermicidní látky a samice může volit spermie určitého samce – bezpečně to bylo prokázáno pomocí radioizotopového značení u štěnic, které mají zvláštní spermatolytické orgány, v nichž selektují pro ně „nevhodné“ spermie a využívají je jako potravu. Na tomto příkladu lze také dobře ukázat, že ke kompetici samců může docházet až uvnitř samice a jen těžko lze rozlišit samčí kompetici a samičí volbu. Metaforicky lze hovořit o „vnitřních námluvách“.
 Traumatická inseminace může vést ke skutečně bizarním situacím. Tak např. africké netopýří štěnice z podčeledi Afrocimicinae mají bisexuálně zaměřené samce, kteří kopulují jak se samicemi, tak vzájemně mezi sebou. Řeknete si, nic nového v šeru netopýřích úkrytů. Je to ale jediný známý případ, kdy je samčí homosexualita do té míry vázaná, že pro homosexuální traumatické kopulace existují u samců specifické orgány, podobně utvářené a umístěné jako příslušné orgány samic. Zatím jediným přijatelným vysvětlením je druhotná potravní funkce inseminace samců. Tyto štěnice se pravidelně ocitají v hladovém stresu a samci, kteří jsou pasivněji „gay“, získávají nadbytečné množství spermatozoální potravy, která jim umožňuje déle přežít, což by mohlo i zvýšit jejich možnosti kopulace se samicemi. Podrobnosti ze života v přírodě však známy nejsou. Má-li někdo z čtenářů lepší hypotézu, bude vítána.
Traumatická inseminace může vést ke skutečně bizarním situacím. Tak např. africké netopýří štěnice z podčeledi Afrocimicinae mají bisexuálně zaměřené samce, kteří kopulují jak se samicemi, tak vzájemně mezi sebou. Řeknete si, nic nového v šeru netopýřích úkrytů. Je to ale jediný známý případ, kdy je samčí homosexualita do té míry vázaná, že pro homosexuální traumatické kopulace existují u samců specifické orgány, podobně utvářené a umístěné jako příslušné orgány samic. Zatím jediným přijatelným vysvětlením je druhotná potravní funkce inseminace samců. Tyto štěnice se pravidelně ocitají v hladovém stresu a samci, kteří jsou pasivněji „gay“, získávají nadbytečné množství spermatozoální potravy, která jim umožňuje déle přežít, což by mohlo i zvýšit jejich možnosti kopulace se samicemi. Podrobnosti ze života v přírodě však známy nejsou. Má-li někdo z čtenářů lepší hypotézu, bude vítána.
 Aby situace nebyla tak jednoduchá: existují i jevy, které jsem kdysi shrnul pod termín androtraumatická inseminace: Aby samec mohl inseminovat samici, musí si ublížit. Nejde o masochistickou situaci, ale o pravidelný jev. Např. většina pavouků při transportu spermatoforu do semenné schránky (spermatéky) samice si musí odlomit špičku sekundárních genitálií (specializovaného makadla). U včely medonosné si při kopulaci samec utrhne konec obrovského falu a potenciální inseminační následníci jej pak s námahou odstraňují.
Aby situace nebyla tak jednoduchá: existují i jevy, které jsem kdysi shrnul pod termín androtraumatická inseminace: Aby samec mohl inseminovat samici, musí si ublížit. Nejde o masochistickou situaci, ale o pravidelný jev. Např. většina pavouků při transportu spermatoforu do semenné schránky (spermatéky) samice si musí odlomit špičku sekundárních genitálií (specializovaného makadla). U včely medonosné si při kopulaci samec utrhne konec obrovského falu a potenciální inseminační následníci jej pak s námahou odstraňují.  Samci ploštic z podčeledi Phallopiratinae čeledi Enicocephalidae nemají vůbec pohlavní otvor a pro inseminaci si musí odlomit vrchol ostrého falu. Panictví je u samců těchto skupin i morfologicky zřejmé a u některých skupin nejsou samci promiskuitní.
Samci ploštic z podčeledi Phallopiratinae čeledi Enicocephalidae nemají vůbec pohlavní otvor a pro inseminaci si musí odlomit vrchol ostrého falu. Panictví je u samců těchto skupin i morfologicky zřejmé a u některých skupin nejsou samci promiskuitní.
Argumenty proti hypotéze samičí volby
Existují i skupiny – zcela výjimečně, ale přesto existují – u nichž jsou velké mezidruhové rozdíly nikoli u samčích, ale u samičích genitálií. Samci mají naopak genitálie značně monotónní. Lze to vysvětlit tak, že samice nerozpoznávají samce primárně podle genitálních stimulů, ale třeba podle mechanického či zrakového kontaktu, nebo se samci příbuzných druhů mohou výrazně odlišovat ekologicky či etologicky. Samec může např. vysílat různé vibrace, které způsobí, že připravenou samici stimulují i jinak zdánlivě fádní samčí genitálie. Samci některých skupin druhů mají monotónní genitálie i tehdy, kdy bychom očekávali silnou diverzifikaci těchto orgánů. Jsou to většinou skupiny se složitým seznamovacím rituálem, u nichž se kritéria samičího hodnocení posouvají od přímého genitálního kontaktu spíše do předkopulační fáze.
Samci některých skupin druhů mají monotónní genitálie i tehdy, kdy bychom očekávali silnou diverzifikaci těchto orgánů. Jsou to většinou skupiny se složitým seznamovacím rituálem, u nichž se kritéria samičího hodnocení posouvají od přímého genitálního kontaktu spíše do předkopulační fáze.
Teoretickou námitkou může být pokopulační stimulace samice samcem. Tento rituál je možné brát i z lidského hlediska. Samec se ještě po kopulaci snaží „přesvědčit“ samici, že právě on byl tím nejlepším, a chce ji přimět k „použití“ jeho spermií. Postkopulační rituály téměř nebyly studovány. Většinou ani nevíme, kdy během kopulace dochází k inseminaci. Teoreticky je tedy možné, že pokopulační (nebo alespoň poinseminační) chování existuje běžně a my jen nejsme schopni ho rozpoznat.
Závažný argument proti přinesla paleontologie. U hmyzu existuje dokonale zachovaný fosilní materiál (např. z jantaru nebo z anglických rašelinišť) starý i několik set tisíc let a je u něj možné srovnávat genitálie s genitáliemi recentních druhů. A ukázalo se, že u mnoha druhů nedošlo během celé té dlouhé doby k žádné změně. Je to výjimka, o níž nám poskytlo doklad právě oněch pár brouků, nebo jde o univerzálnější jev? Jaký má vlastně diverzifikace genitálií vztah ke speciaci (vzniku nových druhů)? Je její příčinou, nebo důsledkem, anebo snad probíhá právě ve fázi speciace?
Tuto situaci lze ovšem dát do souvislostí s teorií přerušovaných rovnováh (= punktuacionalizmus; J. Gould a N. Eldredge), která předpokládá, že většina období existence druhu je stasigenetická (nedochází k velkým změnám, nejvýše k drobným mikroevolučním adaptacím či driftovým posunům) a že velké změny (v našem případě změny struktur samčích genitálií) probíhají v poměrně krátkém období speciace.
Jako protiargument mohou být chápány i některé bizarní jevy, pro něž zatím neznáme evoluční (někdy dokonce ani funkční) vysvětlení. Tak např. některá zvířátka si genitáliemi šeptají. Jsou známy druhy motýlů a ploštic, které mají v genitáliích stridulační (zvukové vlny produkující) orgány a používají je k epigamní stridulaci. Existují dokonce můry se stridulačním orgánem (u samce), které nemají orgán sluchový. Zjevným smyslem takového ústrojí je rozvibrovat celý genitální systém samice. Řada motýlů z čeledi běláskovitých (Pieridae) má na samčím intromitentním orgánu dokonce oči, či lépe řečeno světločivné orgány.
Možný syntetický pohled
Eberhardova teorie samičí genitální volby zatím nejlépe postihuje situaci existující u mnoha nepříbuzných skupin živočichů. I když vychází z obecné teorie sexuálního výběru (z jiných zájmů samce než samice), přesto neodpovídá plně na kritickou otázku, proč zrovna samci jsou tak proměnliví a svými genitáliemi tak druhově specifičtí.Vyjděme z reálného příkladu, přenesme se do oblastí hypotéz a zkusme učinit závěr. Uvažujme příklad dvou nejprimitivnějších linií ploštic, většinou s bizarně složitými primárními genitáliemi a objímadly samce. Čím je samec složitější, tím je samice jednodušší. Samice těch nejsložitějších samců nemají vnější genitálie vůbec. Jak vysvětlit tento protiklad?
Představme si, že každá skupina s diverzifikovanými samčími genitáliemi měla jako svou primitivní vlastnost promiskuitu samic (u samců je promiskuita většinou samozřejmá) a že monogamie je záležitostí druhotného zjednodušení. Představme si dále, že k tomu, aby mohla samice normálně sexuálně fungovat (tj. nechat se intromitovat, inseminovat a použít příslušné spermie k fertilizaci), potřebuje určitý minimální práh libida. Minimální práh slastného pocitu, kvůli kterému kopuluje a kvůli kterému je v sexuálním kontaktu se samcem. (Uvažujeme momentální etologický a smyslový podnět; konečným rozhodčím však bude pochopitelně dlouhodobá selekční úspěšnost takové reakce.) Samice si však „zvyká“ na stále „kvalitnější“ podněty a prahová hodnota libida se posouvá. Dochází k překotné evoluci. Jak a kde se ale zastaví? Naskýtá se možnost, že existuje určitá maximální prahová hodnota libida. V určité evoluční fázi již pocit přestává být příjemný, slastný a mění se v něco netolerovatelného, např. v bolest nebo jiný stresující pocit (např. u hmyzu nebyly dosud receptory pro bolest zjištěny). To platí pro obě pohlaví. A to by mohl být okamžik, kdy se překotná evoluce zastavuje. Uvažujeme-li tedy o speciaci jako o odštěpení nové populace nebo jako o transformaci populace výchozí, musí s tím být do určité míry spojena i přestavba smyslového systému („naladěného“ na novou stimulaci). Tím by se změnila i hranice maximálně tolerovatelného libida. Nové podněty už by zase mohly být přijímány.

 Obecné vysvětlení nabízejí hypotézy zmíněné v boxu 2 pod čísly 9 a 10. Proč však uvažovat pouze o samičí volbě a libidu jednoho pohlaví? Nejde jen o jednu stranu sexuálně evoluční mince? Obě pohlaví se vyvíjela v koevoluci a vzhledem k jejich různým zájmům můžeme mít za to, že společným příčinným evolučním a selekčně podporovaným faktorem je maximalizace libida obou zprostředkovaná smyslovým tahem. Vzhledem k obecně nevybíravé roli a aktivnější funkci samců jsou u nich evoluční důsledky strukturálně patrné; vzhledem k větší selektivnosti samic (byť nezbytně založené na promiskuitě) se evoluce projevuje spíše na etologické a smyslově funkční rovině (změna preferencí pro přijetí optimálně stimulujícího samce, změna smyslového aparátu spojeného s vnímáním libida, zvýšená funkce orgánů uskutečňujících vnitřní selekci spermií samců a založených na stupni vnímaného libida), zatímco vlastní vnější genitálie zůstávají víceméně uniformní.
Obecné vysvětlení nabízejí hypotézy zmíněné v boxu 2 pod čísly 9 a 10. Proč však uvažovat pouze o samičí volbě a libidu jednoho pohlaví? Nejde jen o jednu stranu sexuálně evoluční mince? Obě pohlaví se vyvíjela v koevoluci a vzhledem k jejich různým zájmům můžeme mít za to, že společným příčinným evolučním a selekčně podporovaným faktorem je maximalizace libida obou zprostředkovaná smyslovým tahem. Vzhledem k obecně nevybíravé roli a aktivnější funkci samců jsou u nich evoluční důsledky strukturálně patrné; vzhledem k větší selektivnosti samic (byť nezbytně založené na promiskuitě) se evoluce projevuje spíše na etologické a smyslově funkční rovině (změna preferencí pro přijetí optimálně stimulujícího samce, změna smyslového aparátu spojeného s vnímáním libida, zvýšená funkce orgánů uskutečňujících vnitřní selekci spermií samců a založených na stupni vnímaného libida), zatímco vlastní vnější genitálie zůstávají víceméně uniformní.
Mnoho situací je protimluvných, ale obecnou tendenci lze identifikovat. Myslím, že nejpodnětnější zobecňující úvahou je nedávná elegantní modelová studie Y. Iwasy a A. Pomianowského (1995), která, byť psaná v matematické řeči a zaměřená na obě pohlaví a sexuální selekci obecně (nikoli na genitálie), ukazuje, že
- samičí preference časem degeneruje (nikoli díky lidsky chápanému individuálnímu zevšednění, ale díky mnohagenerační evoluční nutnosti),
- samice je poté připravena přijmout přednostně samčí novinku – v našem případě pozměněnou samčí genitální strukturu poskytující intenzivnější nebo prostě nové libido. Totéž se vztahuje na samce.
Mnoha témat jsem se dotkl jen mimochodem a zasloužila by si podrobnější rozbor. Je třeba si uvědomit, že problém vysvětlení převládající rozmanitosti genitálií samce a uniformity genitálií samice u většiny skupin živočichů s vnitřním oplozením je jedním ze základních problémů evoluční biologie, fylogenetiky i taxonomie. Genitálie nejsou jen potěšením zvířátek, která je užívají, a taxonomů, kteří je využívají. Jejich stavba, uniformita či rozmanitost a etologie epigamních interakcí jsou i problémem genetiky, teorie speciace, vývojové biologie a mnoha dalších biologických disciplín. Nejde jen o to, co se stane, ale i jak a čím dojde ke kódovanému přenosu informace. Genitálie jej zprostředkují, a proto na ně nelze zapomenout.
Obrázky












Všechno živé na naší Zemi podléhá stálé změně: staré odchází a na jeho místě klíčí nový život. Bez trvalé výměny starého za mladé a nové by život na naší planetě již dávno zmizel. Tento závažný biologický úkol zajišťuje sexuální chování, které je jedním ze základních projevů života...
Chování, které oplodnění umožňuje, nazýváme námluvami nebo tokem, což je nezbytný úvod k páření. Dalšími částmi sexuálních projevů je tvorba párů a chování zajišťující pevnost manželského svazku, který může být dočasný nebo trvalý... Celý rozmnožovací cyklus určují samozřejmě kromě vlastního chování i morfologické, fyziologické, zejména hormonální změny organismu...
U většiny živočichů je nejběžnější polohou páření zadní pozice, samec je obrácen břichem ke hřbetu samice. Primáti, zejména makaci a paviáni (obrázek), se při kopulaci drží chápavýma nohama za lýtka samic. V této poloze se páří i ježci, bodlíni, dikobrazi, u kterých říjná samice přitáhne bodliny či ostny natolik k sobě, aby samci nepůsobila bolest. Lidský způsob kopulace břichy k sobě je znám pravidelně u šimpanze bonobo (Pan paniscus), u velryb, bobrů, sirén, ale i u rejnoků, stonožek, sekáčů a pijavek lékařských. Jeden z nejpozoruhodnějších způsobů páření mají naši nejrychlejší letci - rorýsi. Ti se páří pravidelně v letu. Hroši se páří ve vodě, naproti tomu daleko lépe na vodní způsob života přizpůsobení tuleni a rypouši se páří na suchu...
Délka pohlavního orgánu souvisí se stavbou těla. Sloni mají nápadně dlouhý orgán, podobně i kytovci, lenochodi naopak krátký penis, podobně z primátů i giboni, kdežto šimpanzi vzhledem k naduření okolí vulvy u samice mají penis nápadně dlouhý. Krunýř pásovců zase značně znemožňuje ohebnost těla, a proto mají tito savci jeden z relativně nejdelších pářících orgánů...
Normálně kopulující samci makaka se přidržují na zadních končetinách samice. Izolovaně odchovaní samci jsou sice příchodem samice pohlavně vzrušeni, pokoušejí se však pářit naprosto nenormálním způsobem. U těchto pokusů zůstanou celý život a pářit úspěšným způsobem se již nikdy nenaučí...
Po páření díky vysokým energetickým nárokům jsou samci většiny druhů vyčerpáni, což vedlo k latinskému přísloví Post coitum omnia animalia triste (Po páření jsou všechna zvířata tichá nebo smutná, viz Vesmír 75, 484, 1996/7). Samci myší a potkanů se skulí do klubíčka, opírají se hlavou o zem a upadnou na několik okamžiků do nehybnosti...
O autorovi
Pavel Štys
Doporučujeme

Ničí ozon choleru? 

Jak se člověk stává biologem
