









Morfologie milování
Když profesor Komárek napsal svou knížku „Milování v přírodě“, dostával spoustu dotazů, kdy napíše druhý díl – „Milování doma“...
(úryvek z přednášky)
 Byl první máj, byl lásky čas…
Byl první máj, byl lásky čas…
Zajisté nejen Karel Hynek Mácha, ale každý z nás. Evolučně vzato je však láska v lidském pojetí jen epifenomén milování, pro něž má angličtina výstižný výraz „love-making“.
Obvyklá a očekávaná posloupnost událostí u živočichů s vnitřním oplozením (viz přehled terminologie v boxu 1) je kopulace spojená s intromisí, vyúsťující v inseminaci samice a směřující k její fertilizaci. Vnitřní oplodnění uskutečňují genitálie, které však vykonávají i mnoho dalších funkcí (podrobněji box 3 ve Vesmíru 75, 393, 1996/7).
Zapomeňme nyní na poetickou a altruistickou lásku a věnujme se milování a milovacím strukturám z morfologického a evolučního pohledu. To, o čem budu dále psát, se týká všech živočichů, ale úmyslně opomíjím lásku v kontextu kulturní evoluce, reprodukčně nefunkční milování homosexuální (kromě případu u podčeledi Afrocimicinae – viz druhou část) a zapomínám na hermafrodity. Vše se snažím vidět spíše z hlediska rozmanitosti bezobratlých než lépe známých a člověku bližších obratlovců.
Rozmanitost samčích genitálií
Každý taxonom a morfolog ví, že u mnoha skupin živočichů neexistují takřka žádné somatické rozdíly mezi jednotlivými druhy, přičemž se jejich samčí genitálie mohou nápadně lišit, být až extravagantně utvářené a poskytovat mnoho znaků použitelných v diagnostice.  Samčí genitálie mohou být nejen tvarově až neskutečně bohaté a rozmanité (viz např. obrázek), ale i obrovské,
Samčí genitálie mohou být nejen tvarově až neskutečně bohaté a rozmanité (viz např. obrázek), ale i obrovské, 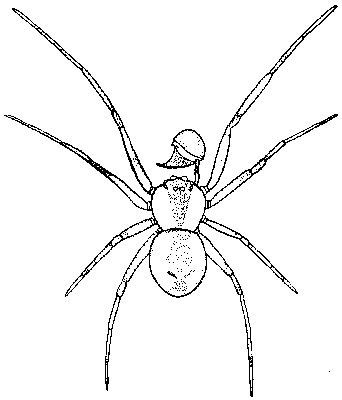 např. u některých ploštic, dvoukřídlých a nematodů jsou ve ztopořeném stavu delší než celé tělo. (Obdobně gigantické mohou být také spermie, např. u některých havajských druhů rodu Drosophila velkých kolem 2,5 mm je spermie s bičíkem 25 mm dlouhá.) Samičí genitálie zůstávají přitom většinou uniformní.
např. u některých ploštic, dvoukřídlých a nematodů jsou ve ztopořeném stavu delší než celé tělo. (Obdobně gigantické mohou být také spermie, např. u některých havajských druhů rodu Drosophila velkých kolem 2,5 mm je spermie s bičíkem 25 mm dlouhá.) Samičí genitálie zůstávají přitom většinou uniformní.
Tyto skutečnosti však téměř nebrala v úvahu teorie evoluce. Až donedávna ji tyto struktury, bez kterých je přenos genetické informace nemožný, nezajímaly. Obecný pohled nenalezneme v žádné učebnici evoluční biologie.
Začněme od Darwina
Roku 1871 formuloval Ch. Darwin svou teorii sexuálního výběru a dal jej do protikladu s výběrem přírodním. Rozeznal dvě základní součásti pohlavního výběru: intrasexuální samčí rivalský souboj o samice a intersexuální samičí volbu samce, který jí nejvíce imponuje a z nějakého důvodu ji přitahuje.  Pohlavní výběr je chápán buď jako součást, nebo jako doplněk přírodního výběru a výrazně ovlivňuje zejména sexuálně dimorfní znaky (projevující se různě u samců a samic). Mohl by vést k tzv. „runaway evolution“ neboli k překotné evoluci, která se nedá zastavit. Samice si vybírají stále atraktivnější vlastnosti samce, až vznikají struktury, které samce samotného handicapují vzhledem k jeho přežití. Aby se takový proces zcela nevymkl kontrole, je – zjednodušeně řečeno – brzděn přírodním výběrem.
Pohlavní výběr je chápán buď jako součást, nebo jako doplněk přírodního výběru a výrazně ovlivňuje zejména sexuálně dimorfní znaky (projevující se různě u samců a samic). Mohl by vést k tzv. „runaway evolution“ neboli k překotné evoluci, která se nedá zastavit. Samice si vybírají stále atraktivnější vlastnosti samce, až vznikají struktury, které samce samotného handicapují vzhledem k jeho přežití. Aby se takový proces zcela nevymkl kontrole, je – zjednodušeně řečeno – brzděn přírodním výběrem.
Je zajímavé, že při zvažování „pohnutek“ samičí volby se vždy braly v úvahu různé atraktivní znaky samců nebo jejich chování, ale opomíjely se vlastní genitálie. Braly se v úvahu jen jako u různých druhů různě vyvinuté orgány sloužící k inseminaci, která posléze vede k oplodnění. Teprve v poslední době se tato tematika stává předmětem zájmu evolučních biologů, kteří formulovali v kontextu pohlavního výběru řadu hypotéz jednak zobecňujících výskyt určitých typů genitálií u živočichů a evoluci jejich rozmanitosti, jednak vysvětlujících jejich funkci, která je složitá a neslouží jen pouhému přenosu spermií.
Typy genitálií živočichů (viz také box 2)
- Do první skupiny patří živočichové, kteří mají genitálie monotónní, morfologicky jednotvárné. Jsou to jednak skupiny druhů s vnějším oplodněním, tedy zejména takové, u nichž živočichové vypouštějí gamety volně do vody, jednak část druhů s vnitřním oplodněním, zejména těch, které mají složité samičí a nikoliv samčí genitálie, nebo těch, které praktikují složité předkopulační rituály (např. ptáci); dále několik málo druhů, kde samec plně převzal samičí sexuální úlohu, a konečně druhy, u nichž samec nekopuluje ani volně nevypouští gamety, ale odkládá spermatofory, které samice vyhledává a sama si je transportuje do těla bez asistence samce.
- Druhá velká skupina, která bez výjimky zahrnuje druhy s vnitřním oplozením, má genitálie velmi často a velmi výrazně druhově specifické. Ovšem jen u samců; samice mají naopak genitálie většinou jednoduché a poskytující jen nevýrazné druhově specifické znaky. Mívají zato velmi komplikovanou vnitřní stavbu a jsou vybaveny složitým smyslovým aparátem.
Situace u samců může být několikerého typu. Běžná a zjevně výchozí je situace, kdy zůstává zachována intromitentní a inseminační funkce primárních genitálií. Často jsou však primární genitálie jednoduché, druhově nespecifické a slouží pouze k vlastní ejakulaci nebo tvorbě spermatoforu. Intromise a inseminace se uskutečňuje za pomoci sekundárních genitálií vzniklých z jiných částí těla, jako je tomu např. u pavouků, kteří používají jako kopulační orgán přeměněná makadla (viz obrázek).
Primární genitálie jsou pak většinou zjednodušené až redukované na pouhý pohlavní otvor. Obdobné situace existují např. u mnoha roztočů, u většiny mnohonožek a u všech vážek. Primární genitálie zde mají pouze ejakulační funkci, vlastní intromisi a inseminaci vykonávají genitálie sekundární, do nichž musí samec svůj spermatofor transportovat. Výjimečná je situace u několika druhů ploštic z čeledi Schizopteridae, u nichž je plně zachován složitý primární, ale pouze ejakulačně fungující genitál napojený na obrovský, ztopořitelný, intromitentní a inseminační genitál sekundární (viz obrázek).
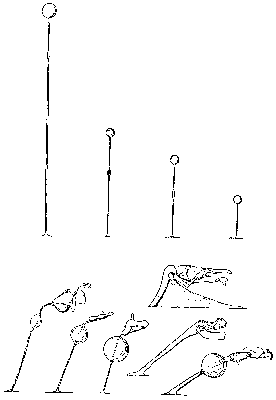 U mnoha samců mnoha druhů z nejrůznějších skupin živočichů mohou existovat ještě objímadla (angl. „claspers“) – klíšťkovité orgány buď tvořící součást primárních nebo sekundárních genitálií, nebo umístěné mimo vlastní genitální oblast, nebo tvořené specificky přeměněnými obvyklými orgány, jako jsou tykadla, nohy, křídla atp. (viz obrázek). Původní představa byla, že jejich funkcí je pouhá fixace samce na samici během kopulačního aktu, nebo rozvírání samičích genitálií pro usnadnění příjmu samčího intromitentního orgánu. Téměř bez výjimky však jsou objímadla opatřena bohatým smyslovým aparátem a dnes převládá myšlenka, že kromě občasné a očividné mechanické funkce jde o orgány zvyšující samčí i samičí libido.
U mnoha samců mnoha druhů z nejrůznějších skupin živočichů mohou existovat ještě objímadla (angl. „claspers“) – klíšťkovité orgány buď tvořící součást primárních nebo sekundárních genitálií, nebo umístěné mimo vlastní genitální oblast, nebo tvořené specificky přeměněnými obvyklými orgány, jako jsou tykadla, nohy, křídla atp. (viz obrázek). Původní představa byla, že jejich funkcí je pouhá fixace samce na samici během kopulačního aktu, nebo rozvírání samičích genitálií pro usnadnění příjmu samčího intromitentního orgánu. Téměř bez výjimky však jsou objímadla opatřena bohatým smyslovým aparátem a dnes převládá myšlenka, že kromě občasné a očividné mechanické funkce jde o orgány zvyšující samčí i samičí libido.
V jistém smyslu jsou sekundárními genitáliemi i specificky druhově utvářené spermatofory, odkládané samcem na substrát a vpravované samicí, popřípadě za spolupráce většinou negenitálních tělních partií samce do vulvy.  Z hlediska následných úvah je pozoruhodná situace u štírků (Pseudoscorpionida), u nichž složité a druhově specifické spermatofory (viz obrázek) vytvářejí samci pouze těch druhů, které mají předinseminační rituál a u nichž skutečně dochází k sexuálnímu kontaktu obou pohlaví, kdy samec většinou asistuje při transportu spermatoforu do těla samice. Ve výčtu zvláštních situací bychom mohli pokračovat do nekonečna, např. někteří chvostoskoci (Collembola) ze skupiny Symphypleona mají dlouhou předkopulační fázi, kdy samice nosí samce přichyceného jeho pozměněnými tykadly (= objímadly), někdy i přehozeného na záda jako pytel brambor. Samec pak sází spermatofory na její hřbet a později, po uvolnění objetí, je přenáší do její vulvy svými ústními orgány.
Z hlediska následných úvah je pozoruhodná situace u štírků (Pseudoscorpionida), u nichž složité a druhově specifické spermatofory (viz obrázek) vytvářejí samci pouze těch druhů, které mají předinseminační rituál a u nichž skutečně dochází k sexuálnímu kontaktu obou pohlaví, kdy samec většinou asistuje při transportu spermatoforu do těla samice. Ve výčtu zvláštních situací bychom mohli pokračovat do nekonečna, např. někteří chvostoskoci (Collembola) ze skupiny Symphypleona mají dlouhou předkopulační fázi, kdy samice nosí samce přichyceného jeho pozměněnými tykadly (= objímadly), někdy i přehozeného na záda jako pytel brambor. Samec pak sází spermatofory na její hřbet a později, po uvolnění objetí, je přenáší do její vulvy svými ústními orgány.
Kuriózní je případ mořských členovců nohatek (Pycnogonida nebo Pantopoda): oplození je vnější, obě pohlaví jsou v podstatě bez genitálií, samec oplozuje vajíčka tak, jak vycházejí z vulvy samice, a okamžitě je přemísťuje na své ovigery (specializovaný pár nohou vlastní pouze samcům) srovnatelné se sekundárními genitáliemi. A není vyloučeno, že vykonávají vzhledem k samici i stimulační funkci.
Intromitentní orgány mají u živočichů pouze samci a k přenosu vajíček či oocytů do těla samce nedochází. Přesto existují dvě výjimky. Jednou z nich jsou ryby z čeledí Hippocampidae (mořští koníčci – viz obrázek a obrázky obrázek, obrázek a obrázek) a Syngnathidae (mořské jehly). Zde samec převzal téměř plně samičí investiční roli. Samice svým intromitentním orgánem přenáší vajíčka do plodové dutiny samce, v ní dochází k jejich oplodnění a dalšímu vývoji (u některých druhů i k pseudoplacentaci). Druhou, analogickou a přitom zcela odlišnou situaci si vyvinuli drobní obojživelní brouci z čeledi Helodidae. Zde sice dochází k obvyklé intromisi samčích primárních genitálií do těla samice, ale ta je vybavena i svým intromitentním orgánem (prehensorem), kterým při kopulaci vniká do genitálií samce, odebírá z nich spermatofor a sama se inseminuje.
A zde se dostáváme k prvním základním otázkám. Proč jsou u druhů s vnitřním oplodněním samčí genitálie tak složité a většinou i druhově specifické?
Proč vznikla celá široká paleta nejrůznějších a nejrůzněji utvářených sekundárních genitálií? Jsou natolik rozšířené a vznikly v průběhu evoluce tolikrát, že je jen těžko možné uplatňovat u každé skupiny jedinečný princip. Obecnou příčinou by mohla být termoregulace: nutnost dočasného uložení spermií na teplotně vhodném místě.
Pravděpodobnějším vysvětlením je ale vliv slasti, libida. Nejlépe nám to ukáže příklad známý u samců pavouků. Ti si před ejakulací z jednoduchých primárních genitálií staví masturbační síť. O ni se třou zadečkem a vypouštějí spermatofor, překládají ho do kopulačních makadel (sekundárních genitálií) a těmi teprve inseminují samici. Protože citlivé topořitelné struktury jsou nejen na primárním genitálu, ale i na makadlech, samčí libido se vlastně zdvojnásobí. Ale to se již ocitáme v oblasti hypotéz.
Teoretické východisko
Nebudeme zde probírat všechny komplikace teorie pohlavního výběru; článek je o genitáliích. Alespoň některá východiska, která relativně nedávno formulovali teoretikové sexuální selekce (např. W. D. Hamilton, G. C. Williams, R. L. Trivers a M. Andersson), bychom si však měli uvědomit. Zájmy samce a zájmy samice jsou totiž již ve svých základech odlišné. Samice většinou investuje více do tvorby gamet a případné výchovy potomstva. Je proto jejím prvořadým zájmem, aby byla vajíčka oplodněna kvalitním samcem – samcem s kvalitními geny. Jejím reprodukčním zájmům většinou neposlouží, když bude oplodněna více samci (spermie jednoho samce naprosto postačují pro veškeré potomstvo samice).Potřebou samce je naopak co nejvíce rozmístit – uplatnit – své geny. Jeho cílem je proto inseminovat a fertilizovat co největší počet samic a zároveň uplatnit paternitu čili znemožnit kopulaci, inseminaci a fertilizaci dalším samcům. Oba požadavky mohou být v rozporu: známý je příklad samce ploštice Nezara viridula (kněžicovití, Pentatomidae), který, aby si zajistil paternitu, dokáže prodloužit kopulaci se samicí až na 80 dnů.
Méně investující pohlaví, to znamená samci, jsou v sexuální kompetici s vlastním pohlavím vzhledem k potenciálnímu oplození dostupných samic. To je zjevně důvod, proč mají intromitentní orgány právě samci a proč je nemají samice (s výše zmíněnou výjimkou). Samci jsou prostě méně vybíraví a jejich rozmnožovací úspěšnost je dána dostupností samic. Dalo by se očekávat, že samice za obvyklých okolností nepotřebují větší počet kopulací spojených s inseminací. Ukážeme si dále, že tomu tak není.
Hypotézy vysvětlující diverzitu samčích genitálií
Jakkoli zde není prostor pro probírání podrobností všech hypotéz a argumentů pro a proti, zmíním se v poměrně úplném přehledu (viz box 4 v druhé části) alespoň o jejich základních východiscích. Mimochodem, všechny ucelené hypotézy jsou nedávného data; do oblasti klasiky patří pouze „zámek a klíč“.
Hypotézy nevycházející primárně ze samičí volby
 Už v roce 1844 formuloval francouzský anatom Léon Dufour ve svém díle „Anatomie générale des diptres“ hypotézu „zámku a klíče“ (známou běžně jako „lock and key hypothesis“), která dominovala v přemítání o diverzitě genitálií (zejména u hmyzu) až donedávna. Dufour se domníval, že druhově specifické genitálie se vyvinuly tak, že znemožňovaly mezidruhovou hybridizaci, čili, v dnešní terminologii, účinkují jako součást RIM (reprodukčně izolačních mechanizmů). Samičí genitálie jako poměrně jednoduchý „zámek“, samčí jako většinou komplikovanější „klíč“, a jen mechanický soulad obou umožňuje úspěšnou kopulaci a inseminaci. Jakkoli je u mnoha druhů mechanický soulad samčích a samičích struktur rafinovaně dokonalý a jakkoli je jasné, že z mechanických důvodů někdy mezidruhovou kopulaci nelze uskutečnit, tento předpoklad jako obecná hypotéza neobstojí. U většiny příbuzných druhů nejsou rozdíly ve struktuře genitálií tak velké, aby mohly zabránit mezidruhové kopulaci, zejména i proto, že samičí genitálie bývají často měkké, roztažitelné a schopné přijmout řadu typů samčích genitálií. Kopulaci zabraňují daleko častěji různé etologické předkopulační mechanizmy, založené zejména na smyslových podnětech. Zcela odlišné samčí genitálie mívají i druhy žijící po stamiliony let v izolaci (např. ostrovní, jeskynní). Nejsložitější a druhově odlišné spermatofory mívají právě ty druhy, u nichž je mezidruhové setkání nepravděpodobné. A např. u ploštěnek, zvláště u těch, které praktikují traumatickou inseminaci (viz níže), může samec kopulovat prakticky s kteroukoli samicí jiného druhu. Ani nedávné experimenty u denních motýlů (např. A. M. Shapiro) teorii „zámku a klíče“ nepodpořily.
Už v roce 1844 formuloval francouzský anatom Léon Dufour ve svém díle „Anatomie générale des diptres“ hypotézu „zámku a klíče“ (známou běžně jako „lock and key hypothesis“), která dominovala v přemítání o diverzitě genitálií (zejména u hmyzu) až donedávna. Dufour se domníval, že druhově specifické genitálie se vyvinuly tak, že znemožňovaly mezidruhovou hybridizaci, čili, v dnešní terminologii, účinkují jako součást RIM (reprodukčně izolačních mechanizmů). Samičí genitálie jako poměrně jednoduchý „zámek“, samčí jako většinou komplikovanější „klíč“, a jen mechanický soulad obou umožňuje úspěšnou kopulaci a inseminaci. Jakkoli je u mnoha druhů mechanický soulad samčích a samičích struktur rafinovaně dokonalý a jakkoli je jasné, že z mechanických důvodů někdy mezidruhovou kopulaci nelze uskutečnit, tento předpoklad jako obecná hypotéza neobstojí. U většiny příbuzných druhů nejsou rozdíly ve struktuře genitálií tak velké, aby mohly zabránit mezidruhové kopulaci, zejména i proto, že samičí genitálie bývají často měkké, roztažitelné a schopné přijmout řadu typů samčích genitálií. Kopulaci zabraňují daleko častěji různé etologické předkopulační mechanizmy, založené zejména na smyslových podnětech. Zcela odlišné samčí genitálie mívají i druhy žijící po stamiliony let v izolaci (např. ostrovní, jeskynní). Nejsložitější a druhově odlišné spermatofory mívají právě ty druhy, u nichž je mezidruhové setkání nepravděpodobné. A např. u ploštěnek, zvláště u těch, které praktikují traumatickou inseminaci (viz níže), může samec kopulovat prakticky s kteroukoli samicí jiného druhu. Ani nedávné experimenty u denních motýlů (např. A. M. Shapiro) teorii „zámku a klíče“ nepodpořily.
 Obdobou Dufourovy hypotézy, zdánlivě jen druhou stranou téže mince, ale z hlediska evoluční kauzality velmi odlišnou představou, je názor H. E. H. Petersona, že takto to v přírodě skutečně funguje, ale že „zámek a klíč“ jsou součástí mechanizmů souborně označovaných jako SMRS (akronym pro „specific mate recognition system“), které umožňují rozpoznat druhově totožného rozmnožovacího partnera a úspěšně sladit všechny vnitrodruhové rozmnožovací aktivity. Ani v této interpretaci mechanistická představa zámku a klíče neplatí, ale jak uvidíme dále, má tato hypotéza mnohem racionálnější základ.
Obdobou Dufourovy hypotézy, zdánlivě jen druhou stranou téže mince, ale z hlediska evoluční kauzality velmi odlišnou představou, je názor H. E. H. Petersona, že takto to v přírodě skutečně funguje, ale že „zámek a klíč“ jsou součástí mechanizmů souborně označovaných jako SMRS (akronym pro „specific mate recognition system“), které umožňují rozpoznat druhově totožného rozmnožovacího partnera a úspěšně sladit všechny vnitrodruhové rozmnožovací aktivity. Ani v této interpretaci mechanistická představa zámku a klíče neplatí, ale jak uvidíme dále, má tato hypotéza mnohem racionálnější základ.
 E. Mayr (1963) E. N. Arnold (1973) se domnívali, že obrovská diverzita genitálií je jen vedlejším produktem pleiotropicky účinkujících genů. Dle Mayra s genitálním efektem původně neutrálním, dle Arnolda vytvářejícím preadaptace pro „zámek a klíč“. Jde zřejmě o mylně formulované hypotézy. Proč by měly být vedlejším efektem postihovány právě genitálie, proč právě samčí, a proč často genitálie sekundární, když primární zůstávají beze změn?
E. Mayr (1963) E. N. Arnold (1973) se domnívali, že obrovská diverzita genitálií je jen vedlejším produktem pleiotropicky účinkujících genů. Dle Mayra s genitálním efektem původně neutrálním, dle Arnolda vytvářejícím preadaptace pro „zámek a klíč“. Jde zřejmě o mylně formulované hypotézy. Proč by měly být vedlejším efektem postihovány právě genitálie, proč právě samčí, a proč často genitálie sekundární, když primární zůstávají beze změn?
Poněkud podnětnější je teorie mechanického konfliktu zájmů, kterou vyslovil americký etolog J. E. Lloyd (1979). V podstatě předpokládá koevoluční závod mezi samci a samicemi. Samci mají jiné zájmy než samice, a proto mezi změnami u samic a změnami u samců neustále dochází ke strukturálnímu vyrovnávání. Ve skutečnosti tomu tak být nemůže: genitálie samce a genitálie 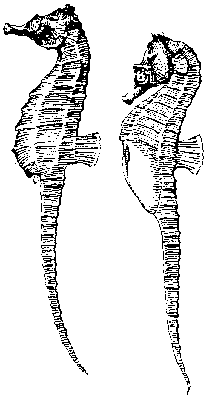 samice se nevyvíjejí stejným tempem a velmi často mezi nimi neexistuje morfologický soulad.
samice se nevyvíjejí stejným tempem a velmi často mezi nimi neexistuje morfologický soulad. 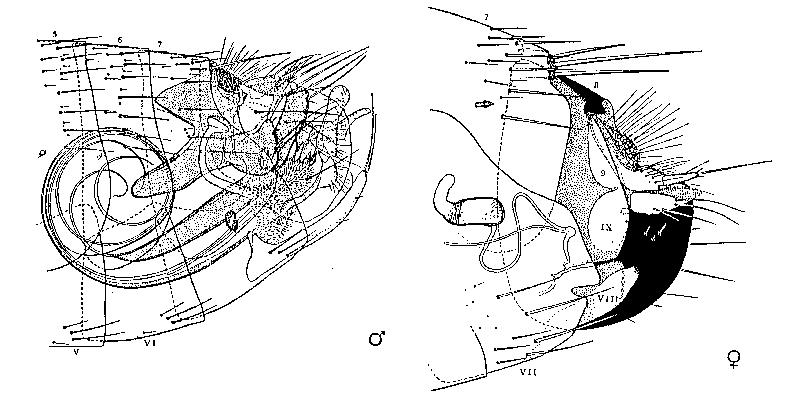 Dle Lloydovy hypotézy se samec i samice snaží mít co největší možnost manipulace s druhým pohlavím. Z hlediska teorie pohlavního výběru argumentují proti této hypotéze skutečnosti, že pro mnohé samice je neschopnost překonat samčí manipulaci selekčně výhodná, že samice nikdy nemají mechanizmy antagonistické k funkci samčích objímadel a že genitálie samců jsou u mnoha druhů neschopny manipulace se samicí.
Dle Lloydovy hypotézy se samec i samice snaží mít co největší možnost manipulace s druhým pohlavím. Z hlediska teorie pohlavního výběru argumentují proti této hypotéze skutečnosti, že pro mnohé samice je neschopnost překonat samčí manipulaci selekčně výhodná, že samice nikdy nemají mechanizmy antagonistické k funkci samčích objímadel a že genitálie samců jsou u mnoha druhů neschopny manipulace se samicí.
Hypotézy založené na samičí volbě (intersexuální selekci)
Hypotézy, které budou zmíněny v boxu 4 (druhá část, Vesmír 75, 393, 1996/7) pod čísly 6 – 8 se prolínají a nevylučují. Všechny jsou založeny na principu samičí volby uskutečňované často dle kvality prožité kopulace, jinými slovy dle kvality struktur samčího partnera uskutečňujících kopulaci (ať již jde o primární nebo sekundární genitálie nebo o objímadla) a dle dovednosti je používat (mluvíme samozřejmě o geneticky podmíněných vlastnostech a neuvažujeme je v lamarckistickém kontextu).  Samice dovolí inseminaci tomu samci a rozhodne o fertilizaci spermiemi takového samce, který jí přivodil maximálně snesitelné libido (viz závěr, 2. část) nebo který dokázal udržet svou paternitu. Což může (a nemusí) být jedno a totéž. R. Thornhill a J. Alcock (1983) předpokládali víceméně automatické přijetí genitálně nejzdatnějšího samce a hlavní význam přisuzovali samčímu soupeření o fertilizaci. W. G. Eberhard, který se této problematice věnoval nejpodrobněji a nejdůsledněji – např. 1985: Sexual Selection and Animal Genitalia – uvažuje spíše o významu aktivního samičího výběru mezi různě intenzivně (či stejně intenzivně, ale jinak) stimulujícími samci. Samec se snaží o úspěšnou propagandu svých genitálních vlastností, samice si vybírá partnera dle jeho genitálních kvalit, a celou sekvenci událostí lze metaforizovat jako „smyslový tah“ (angl. „sensory drive“), kdy samice dává přednost situacím zvyšující její libido. Samec s novou genitální vlastností může účinkovat jako „smyslová past“, kdy jeho neobvyklá struktura stimuluje preadaptované, již existující, ale v sexuálním kontextu dosud nefungující samičí receptory schopné zvyšovat libido. Za předpokladu promiskuity samice má větší naději inseminovat samec, který více stimuluje, a samice může preferovat i jeho spermie. S tím může souviset opět v genitálním kontextu chápaný jev:výhoda (přednost) vzácného samce – promiskuitní samice dává přednost nové vlastnosti, s kterou se dosud nesetkala; prostě proto, že neobvyklost samčí vlastnosti zvyšuje samičí libido bez ohledu na adaptivní hodnotu příslušné samčí vlastnosti.
Samice dovolí inseminaci tomu samci a rozhodne o fertilizaci spermiemi takového samce, který jí přivodil maximálně snesitelné libido (viz závěr, 2. část) nebo který dokázal udržet svou paternitu. Což může (a nemusí) být jedno a totéž. R. Thornhill a J. Alcock (1983) předpokládali víceméně automatické přijetí genitálně nejzdatnějšího samce a hlavní význam přisuzovali samčímu soupeření o fertilizaci. W. G. Eberhard, který se této problematice věnoval nejpodrobněji a nejdůsledněji – např. 1985: Sexual Selection and Animal Genitalia – uvažuje spíše o významu aktivního samičího výběru mezi různě intenzivně (či stejně intenzivně, ale jinak) stimulujícími samci. Samec se snaží o úspěšnou propagandu svých genitálních vlastností, samice si vybírá partnera dle jeho genitálních kvalit, a celou sekvenci událostí lze metaforizovat jako „smyslový tah“ (angl. „sensory drive“), kdy samice dává přednost situacím zvyšující její libido. Samec s novou genitální vlastností může účinkovat jako „smyslová past“, kdy jeho neobvyklá struktura stimuluje preadaptované, již existující, ale v sexuálním kontextu dosud nefungující samičí receptory schopné zvyšovat libido. Za předpokladu promiskuity samice má větší naději inseminovat samec, který více stimuluje, a samice může preferovat i jeho spermie. S tím může souviset opět v genitálním kontextu chápaný jev:výhoda (přednost) vzácného samce – promiskuitní samice dává přednost nové vlastnosti, s kterou se dosud nesetkala; prostě proto, že neobvyklost samčí vlastnosti zvyšuje samičí libido bez ohledu na adaptivní hodnotu příslušné samčí vlastnosti.
Hypotéza samičí volby vysvětluje řadu skutečností:
- Především to, že samčí intromitentní orgány se vyvíjejí rychleji než orgány jiné. Potvrzují to i nálezy mnoha druhových komplexů (třeba až téměř kryptických druhů), které se somaticky vůbec neliší, ale samčí genitálie mají odlišeny ostře.
- Vysvětluje také, proč u druhů se složitými a specifickými druhotnými genitáliemi zůstaly primární genitálie jednoduché nebo se zjednodušily.
- Vysvětluje i to, proč jsou genitálie druhů s vnějším oplozením bez výjimky jednoduché. Nikdy se nezesložiťují, protože na ně nepůsobí samičí volba na základě stimulace.
- Částečně vysvětluje i jistou, i když ne zcela pravidelnou korelaci mezi relativně jednoduchými genitáliemi a složitým chováním při námluvách a složitým systémem epigamních signálů. V těchto případech totiž nastupují jiné vlivy a jiné signály pro samici. Stimulace genitálií ustupuje do pozadí. Ještě více se zde také stírá hranice mezi intrasexuální konkurencí samců (rivalské souboje) a mezi volbou samce samicí. (Genitálie jsou k rivalským soubojům využívány jen zcela výjimečně – např. při rituální agresivní hrozbě u některých primátů.)
- Pomáhá vysvětlit i vznik některých specializovaných negenitálních struktur (objímadel) používaných při kopulaci. Různé druhy hmyzu jsou např. vybaveny nejrozmanitějšími uchopovacími a příchytnými orgány. U samců jsou často vrcholně druhově specifické, každý druh má úplně jiné, ale samicím chybějí – doklad toho, že se samčí, ne sice genitální, ale v sexuálním kontextu používané struktury vyvíjejí na základě samičí volby.
Samčí diverzita může být ovlivněna jistě i mnoha dalšími faktory, např. schopností poskytnout samici genitální cestou nezbytnou doplňkovou výživu (ať už produkcí nadměrného množství spermií nebo semenné tekutiny, vytvářením části spermatoforu, která má jen výživnou funkci [tzv. spermatophylax, např. u kobylek], schopností mechanicky uzavřít samičí vulvu).
(dokončení v následujícím čísle)
Přehled terminologie
ejakulace – vypuštění spermií (a semenné tekutiny) anebo spermatoforu ze samčích primárních genitálií
epigamní – týkající se projevů obou pohlaví v souvislosti s kopulací nebo jinou sexuální interakcí
fertilizace (oplození) – splynutí spermie a oocytu, vznik zygoty
genitálie (samčí) – soubor specializovaných vnějších nebo vychlípitelných struktur zajišťujících kopulaci, ejakulaci a inseminaci, popřípadě další sexuální funkce (viz box 3)
genitálie primární – genitálie s pohlavním otvorem (gonoporem) samce včetně bezprostředně okolních struktur
genitálie sekundární – samčí genitálie bez gonoporu (pohlavního otvoru) a vzdálené od něj; buď striktně genitální struktury, nebo orgány s původně negenitální funkcí a zčásti či zcela v genitálie přeměněné
genitálie ejakulační – genitálie uskutečňující ejakulaci (vždy jde o genitálie primární)
genitálie inseminační – genitálie (primární nebo sekundární) uskutečňující inseminaci
inseminace – přenos spermií nebo spermatoforu do samičích (většinou vnitřních) genitálií, ať už uskutečněný samcem, samicí nebo společně
intromise – vnoření samčích genitálií do těla samice
intromitentní orgán – ta část samčích genitálií, která uskutečňuje intromisi
koevoluce – vzájemně podmíněná evoluce ekologicky či jinak na sebe vázaných taxonů, popřípadě analogická evoluce samců a samic téhož druhu
kopulace – sexuální kontakt obvykle dvou jedinců různého pohlaví téhož druhu směřující k inseminaci (zjednodušeno: neuvažovány hermafroditické a homosexuální kopulace)
libido – (sexuální) slast
objímadlo – samčí struktura (ať již v oblasti primárních nebo sekundárních genitálií nebo mimo ně) fixující nebo objímavě stimulující samici nebo rozevírající její genitálie (nový český termín pro anglické „clasper“)
oocyt – zralá samičí pohlavní buňka (někdy totožná s pojmem vajíčko, to však mívá navíc různé obaly
ovipozice – kladení vajíček
ovulace – uvolňování oocytů (vajíček) ze samičích reprodukčních orgánů za situace, kdy je možná jejich fertilizace
paternita – „otcovství“; skutečnost, že kopulačně investované spermie daného samce skutečně fertilizovaly samici
pleiotropizmus – ovlivnění exprese struktury (nebo její vlastnosti) sadou genů; často spojeno s projevem jiné, funkčně nespojité, a třeba i adaptivně neutrální vlastnosti
preadaptace – funkce, kterou organizmus nebo jeho struktura nevykonává (v daném čase a aktuálním adaptivním kontextu), ale kterou je schopen uskutečnit za změněné adaptivní či environmentální situace – metaforicky: náhodná připravenost na jinou budoucnost
přednost vzácného samce (angl. rare male advantage) – situace, kdy samičí výběr preferuje v sexuálním kontextu samce s neobvyklými (byť třeba z hlediska přírodního výběru neutrálními až maladaptivními) vlastnostmi
pseudokopulace – pokus o kopulaci nebo činnost totožná s kopulací, ale nezavršená funkční inseminací (často přerušená díky zásahu samice)
smyslový tah – tendence (selekčně podmíněná) jednoho nebo dvou pohlaví preferovat sexuální situace zvyšující libido (nový český termín pro angl. sensory drive)
spermatofor – soubor spermií uzavřených ve společném obalu vytvářený samcem před, během nebo po inseminaci, popřípadě samicí v jejích vnitřních cestách, často ukládaný samcem na substrát (mnohdy bez přítomnosti samice) nebo na tělo samice
stasigeneze – fáze vývoje druhu (či vyššího taxonu), kdy nedochází k zásadním evolučním změnám, ale jen k drobným adaptivním úpravám
stridulace – produkce zvukových nebo analogických vln u členovců uskutečňovaná buď vibrací membrán, nebo třením dvou částí těla o sebe (= strigilace)
traumatická inseminace – inseminace uskutečněná zraňujícím proniknutím samčího intromitentního orgánu pokožkou samice, prorážením samičích pohlavních cest nebo jejích specializovaných orgánů (ať už v genitální či negenitální oblasti); spermie jsou injikovány do tělní dutiny samice (popřípadě přes dočasné uložení ve specializované tkáni)
vnější oplodnění – situace, kdy se samčí a samičí gamety setkávají volně v prostředí
vnitřní oplodnění – situace, kdy je inseminace a následná fertilizace dosažena intromisí samčích genitálií do samice nebo transportem spermatoforu (samcem, společně, samicí) do samičích genitálií
vulva – vnější pohlavní otvor samice
O autorovi
Pavel Štys
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
