










Myš domácí
Mezidruhová a mezirasová hybridizace je fenomén, který se stále více dostává do popředí zájmu biologů. Důvodů pro tento zájem je hned několik. Již sama skutečnost, že mezi dvěma populacemi může docházet k vzájemné výměně genů, a přesto si každá uchovává svou genetickou identitu, je z pohledu biologa zajímavá, i když z hlediska striktního pojetí biologického druhu poněkud „nepatřičná“. Podle obecně známé a široce používané definice Ernsta Mayra totiž populace, které při vzájemné hybridizaci dávají ve druhé dceřiné generaci nebo při zpětném křížení vzniknout životaschopnému a plodnému potomstvu, patří ke stejnému druhu. Nicméně existují taxony, mezi nimiž dochází k výměně genetického materiálu, aniž by se stírala jejich individualita, a které jsou všeobecně (tedy i samotnými propagátory biologického konceptu druhu) považovány za plnohodnotné druhy (klasickým příkladem z naší fauny jsou kuňka obecná a kuňka žlutobřichá). 
Hybridní zóny, to jest oblasti, ve kterých se dvě geneticky odlišné populace setkávají, kříží a tvoří potomstvo, lze tudíž chápat jako jakési přírodní laboratoře, poskytující experimentální materiál pro zkoumání podstaty rozdílů mezi vznikajícími druhy, mechanizmů vzniku nových druhů a role, kterou při speciaci hrají náhodné genetické procesy a přírodní výběr.
Mezi modelové objekty sloužící ke studiu mezidruhové či mezirasové hybridizace patří také skupina domácích (tj. většinou žijících v těsné blízkosti lidských sídlišť) myší, kterou bychom mohli nazvat Mus musculus sensu lato. K nejlépe prozkoumaným případům patří křížení M. musculus/M. castaneus na Dálném východě, a především hybridní zóna mezi M. musculus a M. domesticus v Evropě.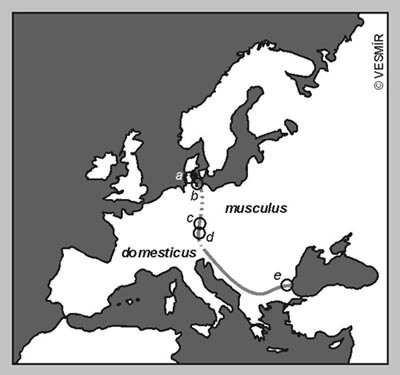
Jak bylo uvedeno v taxonomickém přehledu, areály M. domesticus a M. musculus se v Evropě setkávají podél zhruba 2000 km dlouhé linie, tvořené úzkým pásem hybridních populací. Tato hybridní zóna probíhá napříč Jutským poloostrovem a od pobřeží Baltského moře se táhne až k moři Černému (viz obr. obrázek).
Ačkoli se vzájemný kontakt obou taxonů neomezuje pouze na evropský kontinent, situace v jiných oblastech, např. v Zakavkazí, je zatím známa v mnohem menší míře. Rozsáhlejší výzkum byl dosud prováděn v Dánsku, Bulharsku a v jižním a severním Německu. V poslední době se pozornost začíná upírat rovněž k oblasti západních Čech, kde byl nedávno poprvé prokázán stálý výskyt M. domesticus, o níž se dosud soudilo, že v naší fauně chybí. Předběžné výsledky ukazují, že hybridní zóna mezi oběma druhy, široká 20 – 30 km, protíná ve zhruba poledníkovém směru nejzápadnější cíp našeho státu.
Při zkoumání charakteru hybridní zóny se obvykle sleduje změna vybraných diagnostických znaků odlišujících tyto dva taxony, tzv. markerů, podél transektu (kolmé roviny odhalující vertikální strukturu) vedoucího napříč touto zónou. Nejčastěji se využívají autozomální enzymatické geny s fixovanými alternativními alelami (zpravidla 3–10 lokusů) a znaky morfologické, ale v poslední době se uplatňují i metody molekulární genetiky a cytogenetiky.
Hybridní zóna mezi M. musculus a M. domesticus má pro jednotlivé enzymatické lokusy podobný charakter. To znamená, že gradient, který graficky znázorňuje přechod mezi dvěma diagnostickými alelami, má pro každý lokus zhruba stejný tvar a geografickou polohu, ačkoli u některých lokusů může být v důsledku pronikání cizích alel (introgresi) přes hybridní zónu tento přechod méně ostrý. Zvláštním rysem introgrese je v tomto případě její asymetričnost, způsobená tím, že průnik alel M. domesticus do areálu M. musculus je výraznější než v opačném směru a může dosahovat i do značné vzdálenosti. Kupříkladu alely M. domesticus byly nalezeny v populacích myší v jižních Čechách i v Mostecké pánvi, tedy více než 100 km od předpokládaného centra hybridní zóny.
Chování mitochondriální DNA i morfologických znaků v hybridní zóně je podobné jako chování jaderných (přesněji řečeno autozomálních) genů. Jinými slovy, mezi gradienty pro různé genetické markery – s výjimkou pohlavních chromozomů a některých cytogenetických struktur (viz dále) – nejsou podstatné rozdíly.
Zajímavá situace však byla zaznamenána u skandinávských myší, jejichž jaderné geny jsou typické pro M. musculus, ale jejich mtDNA je typu domesticus. Tento jev, dříve uváděný jako příklad extrémní introgrese (vnesení genů do genomu jiného druhu) mimojaderného genomu, byl teprve v nedávné době vysvětlen na základě zevrubného genetického vyšetření mtDNA i dalších genetických komponent u myší z jižního Švédska, Dánska, Finska a Německa. Ukázalo se, že po obou stranách hybridní zóny ve východním Holštýnsku se v myších populacích nachází mtDNA M. domesticus několika typů, z nichž všechny byly nalezeny rovněž u skandinávských M. musculus, ale nikoli u ostatních populací M. domesticus z jiných částí areálu. Podle autorů studie byla Skandinávie kolonizována malými skupinkami myší, tvořenými vždy několika málo jedinci. Tyto imigrační skupinky pocházely z oblasti zmíněné holštýnské hybridní zóny, odkud se přes poměrně hustou síť ostrovů v Baltském moři dostaly do jižního Švédska a odtud dále na sever. Při jejich dostatečně malé velikosti (teoreticky je možná kolonizace jedinou březí samicí) mohlo dojít k náhodné fixaci mtDNA domesticus v populaci s převážujícími jadernými geny typickými pro M. musculus. Tento jev se většinou označuje odborným termínem „efekt zakladatele“ (founder effect), který vyjadřuje založení nové populace několika málo jedinci, u nichž je díky jejich nízkému počtu vysoká pravděpodobnost častějšího výskytu jinak relativně vzácných genetických variant. V případě mtDNA může být celý proces usnadněn některými specifickými vlastnostmi této molekuly, jako je např. nepřítomnost rekombinace nebo dědičnost téměř výhradně po mateřské linii.
Nápadná shoda struktury a zpravidla i šířky hybridní zóny v různých částech Evropy a pro různé diagnostické znaky vyvolává otázku, jakými faktory je tato koincidence udržována. Poloha zóny vedla některé vědce k domněnce, že její charakter je určován buď klimatickými podmínkami (rozhraní mezi oceánským a kontinentálním podnebím), nebo jinými ekologickými faktory (podobnost s hybridními zónami u jiných druhů živočichů). Je však nepravděpodobné, že by v tomto případě změna vnějších faktorů byla natolik prudká, aby mohla způsobit ostrý přechod jedné formy v druhou. Mnohem pravděpodobnější se zdá hypotéza, že hybridní zóna M. musculus/ M. domesticus je udržována rovnováhou mezi migrací jednotlivých zvířat a selekcí proti hybridním jedincům. Dalo by se tedy říci, že je v určitém „napětí“, a proto je pro tento typ zóny používán termín „tenzní zóna“.
Ačkoli neexistuje žádný přímý doklad o snížené reprodukční způsobilosti (fitness) ani o narušení stability ontogenetického vývoje hybridů, které by byly vyvolány selekcí proti heterozygotním či rekombinantním jedincům, některé nepřímé důkazy nasvědčují tomu, že hybridní genom myší může být skutečně přírodním výběrem znevýhodňován. Např. bylo zjištěno, že myši z hybridní zóny mají prokazatelně vyšší procento střevních parazitů než oba rodičovské druhy a že tento rozdíl je geneticky podmíněn. Navíc přechod pohlavních chromozomů přes hybridní zónu je silně omezen na několik málo kilometrů. Podobný jev byl zjištěn i u některých cytogenetických markerů, jako jsou např. robertsonské dvouramenné chromozomy nebo organizátory jadérek. Tuto hybridní zónu proto někteří autoři považují za bariéru, která chrání genové výbavy proti pronikání cizích alel a zabraňuje nežádoucí rekombinaci nekompatibilních genomů obou druhů.
Odlišná situace existuje na Dálném východě. Areály M. musculus a M. castaneus se zde široce překrývají a mezi oběma taxony dochází k rozsáhlé hybridizaci, jejímž výsledkem je nová forma, tzv. „Mus molossinus“, která se vyskytuje na japonském souostroví, v severovýchodní Číně, Koreji a okrajově pravděpodobně zasahuje i do oblasti ruského Primorje. Analýza geografické struktury mitochondriálních markerů zejména u japonských populací ukázala, že myši se sem dostaly spolu s člověkem nejméně ve dvou kolonizačních vlnách. Jako první osídlila Japonsko M. castaneus, a to z jihovýchodní Asie, za ní následovala M. musculus ze severní Číny. Křížením těchto dvou taxonů vznikla unikátní populace se smíšenou výbavou jaderných genů a geograficky členěnou mozaikou mtDNA obou rodičovských forem: mtDNA typu castaneus na severu a jihu, musculus pak v centrálních oblastech Japonska. Přítomnost dalších dvou typů cytoplazmatického genomu (domesticus, bactrianus) v jižní části souostroví naznačuje původ některých japonských populací domácích myší i z jiných částí světa (obr.
Vzhledem k tomu, že u M. molossinus nebyly nalezeny druhově specifické genetické znaky a samotné japonské populace jsou značně heterogenní, není tato forma obvykle považována za samostatný taxon. Naproti tomu existují určité známky postupné diverzifikace (rozrůzňování), projevující se např. v rozdílné frekvenci alel hemoglobinu nebo velikosti a rozmístění centromerického heterochromatinu.
Ať už bude další evoluční osud M. molossinus jakýkoli, její vznik je z genetického hlediska nesporně zajímavým jevem a spolu s hybridizací M. musculus a M. domesticus v Evropě poskytuje názorný pohled do přírodní kuchyně evoluce. 
Obrázky

Jak již bylo naznačeno v článku o systematickém třídění domácích myší, rozdělení na komenzální a exoantropní formy může být do jisté míry zavádějící, neboť u druhů synantropních mohou některé populace žít po celý rok mimo lidská obydlí a naopak, i u volně žijících druhů může, byť ojediněle, dojít k osídlení budov. Rámcově však toto členění ilustruje rozdíly v ekologické strategii mezi oběma skupinami, a navíc odráží současné představy o fylogenetickém vývoji celého komplexu.
Mus spretus je vázána na oblasti s teplým středomořským klimatem o nadmořské výšce do 1000 m. Obývá plochy pokryté hustou vegetací, např. trávou, bylinami, zemědělskými plodinami nebo křovím. Výzkumy prostorové a sociální organizace některých jihofrancouzských populací ukázaly, že samci tohoto druhu žijí usedle na zarostlých místech a aktivně brání poměrně velké teritorium. Většinou nejsou schopni úspěšně kolonizovat nová, vegetací méně krytá stanoviště. Velmi zajímavý, i když zřejmě zcela mimořádný případ zaznamenal R. D. Sage v jižním Španělsku: M. spretus osídlila obě patra betonové novostavby stojící v pusté krajině mimo městskou zástavbu, zatímco typická obyvatelka budov, M. domesticus, se zde vyskytovala jen v nepatrném počtu.
Mus macedonicus je co do výběru stanoviště poměrně plastickým druhem. V Řecku se vyskytuje na suchých místech s roztroušenými keři a řídkou bylinnou vegetací, na křovinatých mezích, v olivových hájích i na vlhkých, rákosem porostlých stanovištích v okolí jezer, kanálů a jiných vodních toků. V Turecku byla nalezena také ve vegetaci okolo cest a v obilných polích. I přes tuto rozmanitost je M. macedonicus úzce vázána na mediterránní klimatickou zónu a suchým až pouštním oblastem se vyhýbá.
Mus spicilegus je z celé skupiny myší druhem ekologicky nesporně nejzajímavějším. Za svoje jméno (rusky se jí říká kurgančikovaja myš, anglicky hillock mouse, resp. moundbuilding mouse, německy Ährenmaus) vděčí svému zvyku stavět si kopečky z hlíny, které navršuje na zásoby semen (viz obrázek 
Zásobu tvoří nejčastěji obilky a části klasů, semena trav (např. Agropyrum, Setaria, Poa, Sorghum, Digitaria) i jiných rostlin (Centaurea, Zea, Amaranthus, Chenopodium, Polygonum aj.), které myši shromáždí z plochy 10 až 60 m2. Hmotnost těchto zásob se pohybuje kolem 3–6 kg, v některých případech však může dosáhnout až 12 kg! Na pokrytí semen myši používají hlínu z okolí kopečku a z vyhrabaných chodeb. Kopečky bývají nejčastěji na zarostlých okrajích polí a kolem silnic nebo na nekultivovaných, většinou zatravněných pásech mezi obdělávanými poli. Zřídka jsou přímo uprostřed pole, neboť tam bývají ničeny během sklizně nebo orby. Myši začínají se stavbou kopečků již v srpnu (u dočasných staveb dokonce na rozhraní června a července), ale nejintenzivnější činnost vyvíjejí v polovině října; stavební sezona potom může pokračovat až do pozdního podzimu. Vlastní budování jednoho kopečku trvá poměrně krátce: během několika dní až dvou týdnů jsou myši se stavbou hotovy.
V jednom kurgančiku najdeme 5–6, ale někdy i 14 jedinců. V hnízdě myši přečkávají chladné zimní měsíce. Poklesne-li minimální přízemní teplota pod -5 °C během dne a -10 °C v noci, všechny životní funkce se zpomalí a myši upadají do stavu strnulosti, podobného zimnímu spánku. Teplota půdy v hloubce hnízdní komůrky se v té době pohybuje kolem 0 °C. Ačkoli podzemní hnízdo a velké zásoby potravy poskytují poměrně dobré podmínky pro přežití nepříznivého období, úmrtnost myší je vysoká. Jara se nedožije kolem 25–50 % (zpravidla starých) jedinců a během února je už téměř celá populace kurgančiku tvořena pouze nedospělými zvířaty. Je-li zima příliš mírná a bez sněhu, mohou během tohoto období zahynout dokonce všichni obyvatelé kupky. Příčinou zřejmě bývá vysoký výdej energie způsobený zvýšenou aktivitou myší během teplé zimy.
Hustota osídlení v krajině je samozřejmě závislá na úživnosti biotopu, udává se 1 – 20 kopečků na 1 hektar, ale např. v rumunských nížinách, v místech vysoké populační hustoty, bylo na ploše jednoho hektaru napočítáno dokonce 60 – 100 kurgančiků. M. spicilegus může obývat stejná pole společně s M. musculus, avšak během zimy se jejich cesty rozdělí: prvně jmenovaný druh přezimuje ve svých kupkách, zatímco domácí M. musculus se stahuje do budov a stohů. Podle některých autorů se do budov stěhují i někteří jedinci M. spicilegus, ale je pravděpodobnější, že v popsaných případech (např. na Slovensku) šlo o chybnou identifikaci M. musculus. Naopak k překvapivému objevu dospěli vědci na Ukrajině: elektroforézou enzymů bylo prokázáno, že v jednom kurgančiku může s M. spicilegus žít společně (byť zřejmě jen dočasně) i M. musculus.
O autorech
Daniel Frynta
Miloš Macholán
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
