










Inzulinem to nekončí
Cukrovka (diabetes mellitus) je chronické onemocnění, jehož hlavním znakem je porušená glukózová rovnováha v těle, která se projevuje zvýšenou hladinou krevního cukru (hyperglykemií). Porozumění procesům kontroly glukózy na buněčné a molekulární úrovni otevírá cesty k novým způsobům léčby.
Hladinu cukru v krvi reguluje hormon inzulin (Vesmír 101, 40, 2022/1). Je produkován pankreatickými beta-buňkami, jež jsou součástí Langerhansových ostrůvků slinivky břišní (pankreatu). Zvýšená hladina cukru v krvi indukuje výlev inzulinu z pankreatických beta-buněk, který následně signalizuje v periferních tkáních (zejména játrech, kosterní svalovině a tuku), že je potřeba odstranit cukr z oběhu vstřebáním do těchto tkání. Cukr je následně přetvořen na zásobní produkty jako glykogen či tuk.
Rozlišujeme dva typy tohoto onemocnění. Cukrovka 1. typu je autoimunitního původu. Imunitní systém reaguje proti samotným beta-buňkám a jejich úbytek následně způsobí nedostatek inzulinu pro regulaci hladiny glukózy. Cukrovka 2. typu v populaci převažuje a je asociována převážně s obezitou a nezdravým životním stylem. Ačkoliv je inzulin beta-buňkami produkován, snižuje se citlivost tkání k inzulinové regulaci (nastává tzv. inzulinová rezistence) a chronicky se zvyšuje hladina krevního cukru. To má za následek sekundární komplikace v podobě poškození tkání, jako například očí (retinopatie), nervů (neuropatie), ledvin (nefropatie) atd. S růstem životní úrovně v posledních desetiletích celosvětově výrazně narůstá prevalence tohoto onemocnění, zejména cukrovky 2. typu.
Převládající léčba aplikací inzulinu (u cukrovky 1. typu), resp. podáváním antidiabetik a dietou (u cukrovky 2. typu) rozhodně není konečným řešením, neboť představuje poměrně invazivní přístup a pacientům přináší mnohé strasti. Díky novým molekulárním technikám, zejména na úrovni jednotlivých buněk, se v poslední době ukazují další možnosti, jak přispět k zachování zdravých beta-buněk, a tím nástup cukrovky včas odvrátit.
Redoxní rovnováha v beta-buňkách
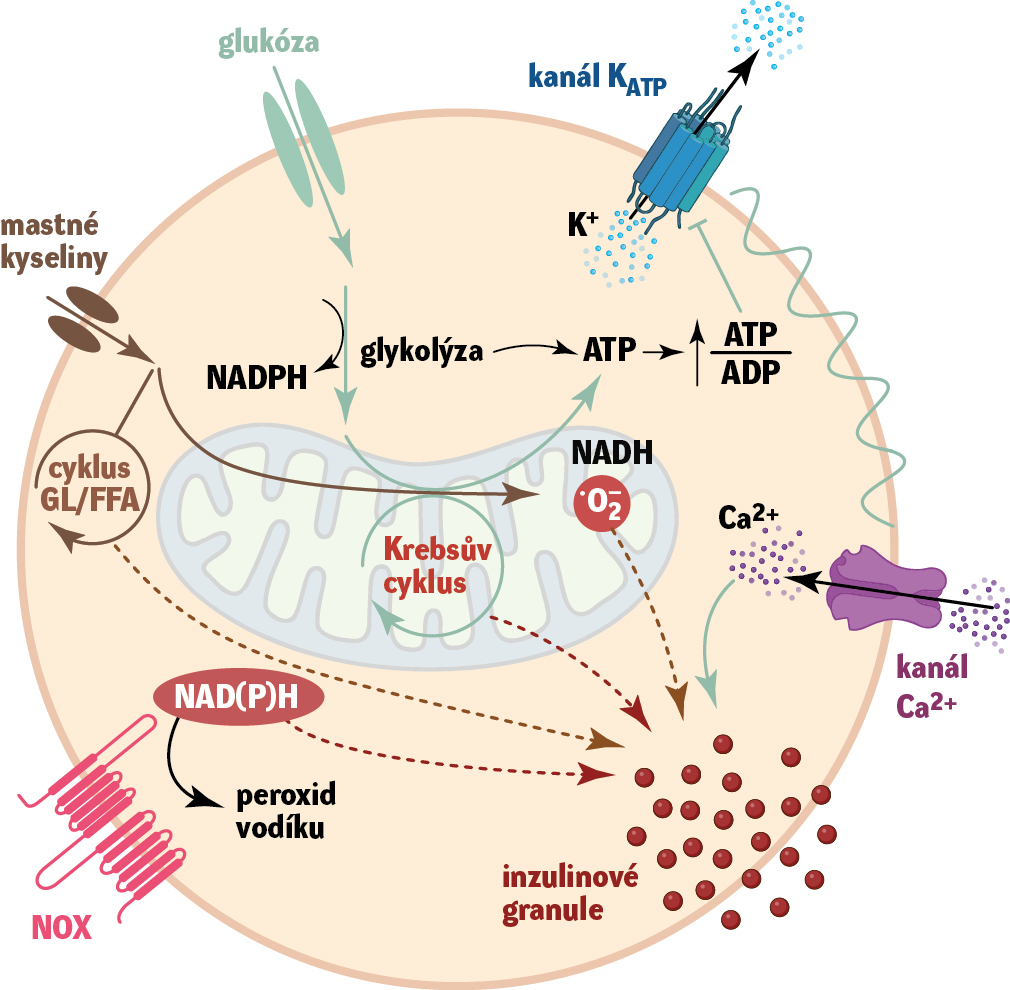
Beta-buňka je majoritní endokrinní buněčný typ, který se nachází ve zdravých Langerhansových neboli pankreatických ostrůvcích slinivky. Již před mnoha lety byl popsán mechanismus výlevu inzulinu jakožto odpovědi na zvýšenou koncentraci glukózy. Zahrnuje metabolickou dráhu zpracování glukózy (glykolýzu), dále oxidativní metabolismus jejích metabolitů v mitochondriích, který vyústí v produkci buněčné energetické molekuly ATP. Ta následně inhibicí kanálu KATP ovlivňuje přenos draslíkových iontů přes plazmatickou membránu buňky. Inhibice kanálu způsobuje depolarizaci plazmatické membrány a následný vtok vápníkových iontů, které umožní vylití inzulinových granulí mimo buňku (obr. 1).
Toto dogma výlevu inzulinu bylo považováno za konečné, a tak další výzkum v oblasti beta-buněk a pankreatických ostrůvků stagnoval. V posledních letech se ale ukazuje, že tento centrální mechanismus výlevu je laděn mnoha dalšími metabolity, které vznikají při zpracování glukózy v beta-buňce a jsou důležité i z hlediska dlouhodobého působení (obr. 1). Jedním takovým metabolitem jsou reaktivní formy kyslíku, zejména peroxid vodíku, které vznikají např. během aktivity mitochondrií při výrobě ATP jako její vedlejší produkt nebo pomocí specifických enzymů během buněčných pochodů. V nízké koncentraci a krátkodobě fungují jako důležité regulátory buněčných pochodů (redoxní signalizace). Zvýšené množství v dlouhodobém horizontu způsobuje jejich nahromadění, tzv. oxidativní stres, který poškozuje buněčné komponenty a je součástí řady patologií včetně cukrovky.
V posledních letech se ukazuje, že právě beta-buňky udržují křehkou redoxní rovnováhu a používají redoxní signalizaci ke správné funkci. Narušení redoxně-oxidační rovnováhy u beta- buněk vede k rozvoji cukrovky. Výzkumem jednoho z enzymů, které tuto rovnováhu v beta-buňkách ovlivňují, NADPH oxidázy 4 (NOX4), se dlouhodobě zabýváme. Zjistili jsme, že během akutní signalizace výlevu inzulinu jakožto odpovědi na zvýšenou přítomnost glukózy dochází k metabolické aktivaci NOX4. Následná produkce peroxidu vodíku spolu s buněčným ATP vedou k efektivnímu výlevu inzulinu a správné funkci beta-buněk (obr. 1). Zároveň ale při chronickém nadbytku živin (a tím i buněčného substrátu, např. glukózy), který je v moderní společnosti běžný (přejídání), může dojít k dlouhodobé aktivaci enzymu NOX4 a jeho zvýšené produkci peroxidu vodíku. Tomu se již beta-buňka v dlouhodobém měřítku neumí bránit. Mění se signalizace, která může vést k aktivaci imunitního systému a zánětu provázejícího cukrovku. Podobně samotné mastné kyseliny, které se při chronickém přejídání uvolňují do krve z tučné stravy v nadbytku, mohou aktivovat produkci kyslíkových radikálů z mitochondrií beta-buněk a redoxní signalizaci dlouhodobě měnit (obr. 1).
Redoxní stav beta-buněk je tedy důležitým hráčem v regulaci fyziologických a patologických pochodů. Jeho účinky závisejí zejména na množství a časové stopě. Jeho detailní výzkum v beta-buňkách může přinést poznatky umožňující včasnou intervenci pomocí specificky cílených antioxidantů. Ta by mohla pomoci oddálit vývoj cukrovky zejména 2. typu.
Buněčná transformace
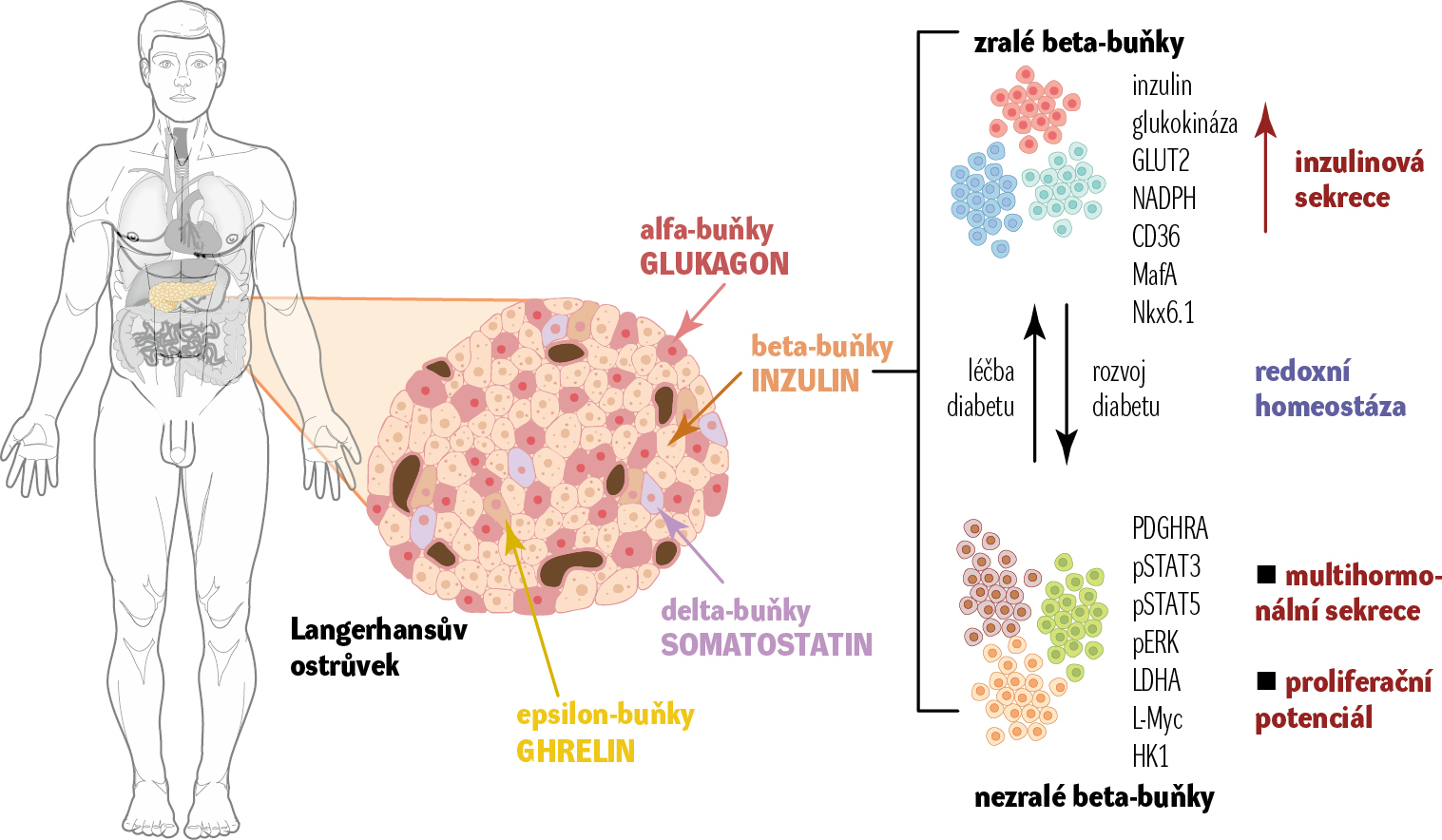
Dalším důležitým poznatkem výzkumu pankreatických beta-buněk je odhalení jejich heterogenity. Metoda RNA sekvenovaní jednotlivých buněk (single cell RNA seq) odhalila, že populace beta-buněk se dělí na buňky metabolicky vyspělé, efektivně odpovídající na výskyt glukózy v okolí výlevem inzulinu, a na buňky nevyzrálé, které si zachovávají spíše proliferační potenciál (obr. 2). V průběhu vývoje diabetu 2. typu se nejprve beta-buňky snaží kompenzovat zvýšenou koncentraci glukózy v krvi. Zvýší produkci a výlev inzulinu (hyperinzulinemie), což vede v dlouhodobém měřítku k jejich vyčerpání (hypoinzulinemie) a v krajních případech ke smrti buněk.
Pro identifikaci beta-buněk v ostrůvcích se používá detekce inzulinu. Pokud na řezech slinivky jeho signál vymizel, např. u vzorků diabetických jedinců, mělo se za to, že došlo k vymizení beta-buněk pomocí programované buněčné smrti (apoptózy). Nyní se ale zjistilo, že metabolicky vyzrálé buňky se vlivem nadměrné produkce inzulinu, která vede k jejich vyčerpání, mohou dediferencovat, aniž by zanikly. Dediferencované buňky jsou často schopné výlevu více hormonů. Kromě inzulinu též glukagonu či somatostatinu, jež jsou za normálních okolností vylučovány výhradně jinými endokrinními buňkami Langerhansových ostrůvků (obr. 2). V případě včasného odeznění metabolického přesycení, např. úbytkem váhy, dietou, se mohu tyto dediferencované beta-buňky opět diferencovat zpět v metabolicky vyzrálé buňky, které obnoví syntézu inzulinu a dalších proteinů potřebných k jejich efektivní odpovědi na zvýšenou glukózu v krvi.
Podobně beta-buňky, které jsou méně vyzrálé, mohou sloužit během života jedince jako „náhradníci“ pro případ zániku zralých beta-buněk vlivem vyhoření. Mohou se pak diferencovat jako jejich náhrada. I zde by svou roli mohla hrát redoxní signalizace a množství reaktivních forem kyslíku. Dokonce se ukázalo, že i další důležité endokrinní buňky Langerhansových ostrůvků, např. alfa buňky, jsou schopné se v krizových situacích adaptace na nepříznivé nutriční podmínky diferencovat v beta-buňky. Alfa buňky za normálních okolností vylučují hormon glukagon, který jakožto protihráč inzulinu za podmínek nízké glukózy v krvi iniciuje naopak uvolňování glukózy z jaterní zásobárny glykogenu.
Této transformační schopnosti jednotlivých subpopulací beta-buněk, ale i dalších endokrinních buněk Langerhansových ostrůvků, by se v budoucnu dalo využít k udržení populace zdravých beta-buněk během života jedince a k zabránění rozvoji diabetu. Transformace endokrinních buněk lze využít i k „laboratorní produkci“ nových zdravých beta- buněk. Ty by pak mohly být transplantovány pacientům, kteří je potřebují.
Ke stažení
 článek ve formátu pdf [947,97 kB]
článek ve formátu pdf [947,97 kB]
O autorovi
Lydie Plecitá Hlavatá

Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
