









Neurónový podklad priestorovej navigácie
 Jednou z veľkých záhad súčasnej vedy je, ako aktivita mozgových buniek reprezentuje prostredie, objekty a vzťahy medzi nimi. Odpovedi na túto všeobecnú otázku sa môžeme priblížiť, ak sa sústredíme na špecifický problém – ako je v neurónovej aktivite mozgu cicavcov reprezentovaný priestor. Mozgová reprezentácia priestoru je pomerne detailne preskúmaná. Boli objavené nervové bunky (neuróny) s fascinujúcimi vlastnosťami, ktorých aktivita odráža priestorové vzťahy: Miestne bunky sú aktívne, ak sa zviera nachádza v určitom mieste prostredia. Bunky smeru hlavy sa aktivujú podľa toho, ktorým smerom je orientovaná hlava zvieraťa, a vlastnosťami pripomínajú kompas. A aktivita mriežkových buniek je organizovaná do pravidelných priestorových obrazcov. Pri pohľade na vlastnosti týchto buniek ťažko odolať predstave, že ich úlohou je reprezentovať priestorové vzťahy a umožňovať navigáciu živočíchov v prostredí. Neurónové dráhy a okruhy, ktoré sa podieľajú na priestorovej navigácii, môžu slúžiť ako modelovýsystém pre štúdium vyšších kognitívnych funkcií, vďaka ktorým my (a ďalšie živočíchy) vytvárame modely okolitého prostredia. Tieto vyššie kognitívne funkcie sú pre mnohých z nás najzaujímavejším aspektom mozgovej činnosti a donedávna boli najmenej preskúmané. Na základe čoraz hlbších a detailnejších poznatkov o neurónovom základe priestorovej navigácie si možno utvoriť obraz o tom, ako nervový systém vytvára mentálne reprezentácie sveta okolo nás.
Jednou z veľkých záhad súčasnej vedy je, ako aktivita mozgových buniek reprezentuje prostredie, objekty a vzťahy medzi nimi. Odpovedi na túto všeobecnú otázku sa môžeme priblížiť, ak sa sústredíme na špecifický problém – ako je v neurónovej aktivite mozgu cicavcov reprezentovaný priestor. Mozgová reprezentácia priestoru je pomerne detailne preskúmaná. Boli objavené nervové bunky (neuróny) s fascinujúcimi vlastnosťami, ktorých aktivita odráža priestorové vzťahy: Miestne bunky sú aktívne, ak sa zviera nachádza v určitom mieste prostredia. Bunky smeru hlavy sa aktivujú podľa toho, ktorým smerom je orientovaná hlava zvieraťa, a vlastnosťami pripomínajú kompas. A aktivita mriežkových buniek je organizovaná do pravidelných priestorových obrazcov. Pri pohľade na vlastnosti týchto buniek ťažko odolať predstave, že ich úlohou je reprezentovať priestorové vzťahy a umožňovať navigáciu živočíchov v prostredí. Neurónové dráhy a okruhy, ktoré sa podieľajú na priestorovej navigácii, môžu slúžiť ako modelovýsystém pre štúdium vyšších kognitívnych funkcií, vďaka ktorým my (a ďalšie živočíchy) vytvárame modely okolitého prostredia. Tieto vyššie kognitívne funkcie sú pre mnohých z nás najzaujímavejším aspektom mozgovej činnosti a donedávna boli najmenej preskúmané. Na základe čoraz hlbších a detailnejších poznatkov o neurónovom základe priestorovej navigácie si možno utvoriť obraz o tom, ako nervový systém vytvára mentálne reprezentácie sveta okolo nás.
Priestorová navigácia
 V živočíšnej ríši existuje mnoho rôznych navigačných stratégií. Príkladom jednoduchej stratégie je pohyb smerom k orientačnému bodu. Takéto správanie môžeme pozorovať pri niektorých druhov hmyzu letiacich za svetlom, ale aj u ľudí, napríklad u turistov sledujúcich značky na stromoch a skalách, keď sa túlaju neznámym terénom.
V živočíšnej ríši existuje mnoho rôznych navigačných stratégií. Príkladom jednoduchej stratégie je pohyb smerom k orientačnému bodu. Takéto správanie môžeme pozorovať pri niektorých druhov hmyzu letiacich za svetlom, ale aj u ľudí, napríklad u turistov sledujúcich značky na stromoch a skalách, keď sa túlaju neznámym terénom.
Ďalším spôsobom navigácie v prostredí je sledovanie súboru inštrukcií, ako to robia ľudia na ceste k cieľu v neznámom meste. Inštrukcie môžu mať napríklad takúto podobu: „Vystúp z metra na Hlavnej ulici a pokračuj po nábreží. Keď sa na pravej strane objaví kostol, zaboč doprava. Hotel bude tretia budova vľavo.“
 Komplikovanejší typ navigácie vyžaduje mentálnu reprezentáciu prostredia, takzvanú kognitívnu mapu. Reprezentáciu založenú na kognitívnej mape, ktorá obsahuje miesta a priestorové vzťahy medzi nimi, používame vtedy, keď sa pohybujeme v prostredí, ktoré dobre poznáme, napríklad v známom meste. Kognitívna mapa prostredia umožňuje využívať skratky či nové, ešte nepoužité cesty.
Komplikovanejší typ navigácie vyžaduje mentálnu reprezentáciu prostredia, takzvanú kognitívnu mapu. Reprezentáciu založenú na kognitívnej mape, ktorá obsahuje miesta a priestorové vzťahy medzi nimi, používame vtedy, keď sa pohybujeme v prostredí, ktoré dobre poznáme, napríklad v známom meste. Kognitívna mapa prostredia umožňuje využívať skratky či nové, ešte nepoužité cesty.
Už v prvej polovici 20. storočia bola vyslovená hypotéza, že aj niektoré zvieratá používajú k navigácii kognitívnu mapu [1]. V príspevku predstavíme nervové bunky a neurónové okruhy, ktoré by mohli takúto kognitívnu mapu vytvárať. Tento neurónový systém reprezentuje aktuálnu polohu zvieraťa, vzťahy medzi rôznymi miestami prostredia a azimut. Neurónové okruhy sa študovali hlavne na hlodavcoch, ale aj na ďalších živočíšnych druhoch, napríklad na opiciach, dokonca aj na netopieroch. Všetky experimentálne výsledky, o ktorých sa zmienime v tomto článku, boli získané pokusmi na laboratórnych potkanoch.
Záznam neurónovej aktivity u voľne sa pohybujúcich zvierat
Naše poznatky o neurónovom podklade priestorovej navigácie vychádzajú z výsledkov experimentov, v ktorých sa nahrávala aktivita jednotlivých neurónov u bdelých, voľne sa pohybujúcich laboratórnych potkanov. Neurónové elektrické výboje (takzvané akčné potenciály) sa zaznamenávali pomocou mikroelektród z veľmi jemných drôtov (25 mikrometrov v priemere). Elektródy boli dlhodobo implantované do mozgu do tesnej blízkosti nervových buniek. Každá z nich zaznamenávala akčné potenciály z niekoľkých neurónov v jej blízkosti. Tvar a amplitúda zaznamenaných akčných potenciálov závisia od polohy a vzdialenosti elektródy od bunky. Akčné potenciály z rôznych neurónov majú preto rôzny tvar, i keď ich zaznamenala tá istá elektróda (obr. 1B). Na základe rôzneho tvaru možno akčné potenciály roztriediť do skupín, tak aby každá skupina obsahovala akčné potenciály z jednej bunky.
 Zväzky mikroelektród sa v narkóze natrvalo voperovali do mozgu a pripevnili ku konektoru na hlave potkana. Zvieratá sa po niekoľkých dňoch zotavili a žili s implantovanými elektródami, často mnoho mesiacov v štandardných laboratórnych podmienkach.
Zväzky mikroelektród sa v narkóze natrvalo voperovali do mozgu a pripevnili ku konektoru na hlave potkana. Zvieratá sa po niekoľkých dňoch zotavili a žili s implantovanými elektródami, často mnoho mesiacov v štandardných laboratórnych podmienkach.
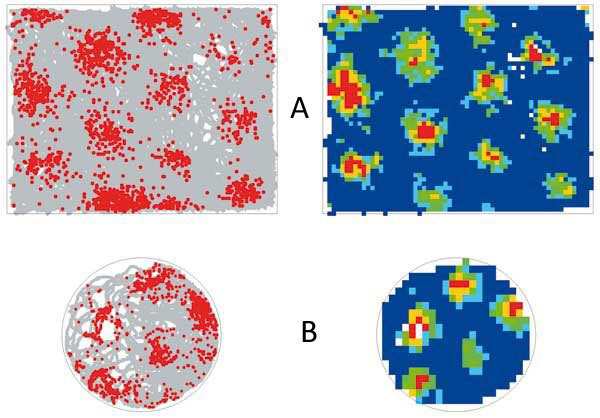
Počas experimentov bol implantát na hlave potkana tenkým káblom pripojený k zosilovačom a ďalším prístrojom potrebným na záznamenávanie aktivity neurónov. Neurónový elektrický signál bol zosílený a filtrovaný, digitalizovaný a nakoniec zaznamenaný na pevnom disku počítača. Uložený signál bol neskôr analyzovaný. Experimenty prebiehali v kruhových arénach s orientačnými bodmi na stenách (obr. 1A). Experimentátor obyčajne pozoroval pokus zo susednej miestnosti na monitore počítača, tam sa nachádzali aj ďalšie prístroje potrebné na zaznamenávanie mozgovej aktivity a správania potkana.
Miestne bunky
Pred vyše tridsiatimi rokmi bolo prvýkrát technicky možné zaznamenávať akčné potenciály z mozgu bdelých, voľne sa pohybujúcich zvierat. Čoskoro objavili na University College v Londýne miestne bunky [2]. Miestne bunky sú neuróny, ktorých aktivita závisí od polohy potkana v priestore. Nachádzajú sa v časti mozgovej kôry zvanej hipokampus. Keď je potkan na určitom mieste, zvanom aktivačné pole, miestna bunka sa aktivuje (vysiela akčné potenciály). Keď sa potkan nachádza mimo aktivačného poľa, bunka je takmer neaktívna. Príklad priestorovej aktivity miestnej bunky je znázornený na obrázku 2A. Táto bunka bola aktívna, keď bol potkan v „severnej“ časti kruhovej arény, v iných častiach arény sa bunka aktivovala len veľmi zriedkavo. Rôzne súčasne nahrávané miestne bunky sa aktivujú v rôznych miestach arény (obr. 2A–D), takže aktivačné polia mnohých miestnych buniek dohromady pokrývajú celý povrch arény, a na základe ich aktivity je možné určiť polohu potkana v prostredí.
Pri detailnom štúdiu miestnych buniek sa ukázalo, že ich vlastnosti v mnohom zodpovedajú tomu, ako subjektívne vnímame priestor. Napríklad, pri orientácii v prostredí sa spoliehame na zmyslové informácie o polohe orientačných bodov. I aktivita miestnych buniek závisí od dôležitých orientačných bodov v prostredí. Ak sa orientačné body premiestia, aktivita miestnych buniek ich nasleduje: aktivačné polia miestnych buniek sa presunú spolu s orientačnými bodmi.
Zmyslové informácie však môžu byť niekedy nespoľahlivé. To isté miesto môže pri rôznych príležitostiach vyzerať rôzne. Napríklad, ak na stenu obývacej izby zavesíme nový obraz, miestnosť bude vyzerať trošku inak, stále ju však budeme považovať za tú istú izbu. Z pokusov vyplynulo, že ani aktivita miestnych buniek obyčajne nezávisí od prítomnosti jednotlivých orientačných bodov, ale skôr od ich celkového usporiadania (celkovej konfigurácie). Odstránenie jedného z nich nezmení priestorovú aktivitu miestnych buniek. Priestorová aktivita miestnych buniek je teda robustná a odolná voči malým zmenám v prostredí, presne tak, ako by sme očakávali od systému, ktorý reprezentuje známe, avšak neustále sa meniace prostredie.
Vráťme sa ešte raz k príkladu s izbou v známom byte. Ak v noci zhasneme svetlá, všetok nábytok, objekty, všetky orientačné body sa stanú neviditeľnými. Napriek tomu stále vieme, kde sme a môžeme sa dokonca poslepiačky, podľa „mapy“ v našej pamäti, pohybovať po miestnosti. Miestne bunky sa pri vypnutých svetlách správajú analogicky. Aj potme si uchovajú priestorovú aktivitu. Aktivujú sa, keď potkan prechádza aktivačnými miestami. V neprítomnosti zrakových orientačných bodov potkan určuje svoju polohu v známom prostredí na základe vlastných pohybov a na základe predošlej skúsenosti s prostredím.
Z pokusov vyplýva, že miestne bunky využívajú zrakové, čuchové, a ďalšie zmyslové informácie, ako aj informácie o pohyboch potkana, avšak ich aktivita nie je úplne závislá od žiadnej jednotlivej zmyslovej modality. Priestorová aktivita miestnych buniek je výsledkom kognitívnych procesov, ktoré kombinujú informácie o vonkajšom prostredí, naučené priestorové vzťahy medzi rôznymi miestami prostredia, ako aj informácie o pohybe potkana, a na základe týchto informácií vytvárajú spoľahlivú reprezentáciu priestoru.
Bunky smeru hlavy
Kompas je veľmi užitočnou pomôckou navigátorov. Hoci to môže znieť prekvapivo, v mozgu potkana sa nachádzajú neuróny, ktoré podobne ako kompas signalizujú azimut. Bunky smeru hlavy, ako ich názov naznačuje, sa aktivujú, ak je hlava potkana orientovaná určitým smerom. Tieto bunky signalizujú rovnaký smer hlavy bez ohľadu na polohu potkana v priestore (obr. 3). Napríklad bunka na obrázku 3 sa aktivuje, keď je hlava potkana orientovaná „severovýchodným“ smerom. Ak je hlava orientovaná na „juh“ či na „západ“, bunka nevysiela akčné potenciály. Rôzne bunky sa aktivujú pri rôznej orientácii hlavy. Bunky smeru hlavy sa nachádzajú v postsubikule a niektorých ďalších častiach mozgovej kôry i v podkôrových oblastiach mozgu, napríklad v talame. Mnohé z týchto mozgových oblastí sú úzko prepojené s hipokampom. Tieto bunky boli objavené na Newyorskej štátnej univerzite v Brooklyne [3].
Na rozdiel od kompasu aktivita buniek smeru hlavy nezávisí od zemského magnetického poľa. Priestorová aktivita týchto buniek vychádza zo zrakových, vestibulárnych a ďalších zmyslových informácií. Dokumentuje to napríklad nasledujúci jednoduchý pokus: ak sú rotované orientačné body okolo kruhovej arény, smer zvýšenej aktivity buniek smeru hlavy rotuje spolu s orientačnými bodmi. Smer hlavy teda nie je detegovaný priamo nejakým špecializovaným zmyslovým orgánom, ale je vypočítaný na základe informácií z rôznych zmyslových modalít. Kľúčovú úlohu zohráva vestibulárny systém vo vnútornom uchu, ktorý deteguje polohu a pohyby hlavy v priestore. Ak je vestibulárny systém poškodený, bunky smeru hlavy stratia svoju priestorovú aktivitu.
Signál o orientácii hlavy v priestore, ktorý je prítomný v aktivite týchto buniek, vychádza zo zmyslových informácií. Tie sú však spracované veľmi komplikovaným a nie do detailov známym spôsobom.
Mriežkové bunky
Ďalšou skupinou neurónov so zaujímavou priestorovou aktivitou sú mriežkové bunky. Tieto neuróny objavili nedávno na univerzite v nórskom Trondheime [4]. Aktivita mriežkových neurónov je v niečom podobná aktivite miestnych buniek. Oboje sú aktívne len v určitých oblastiach prostredia. Každá mriežková bunka má však niekoľko aktivačných polí, na rozdiel od miestnych buniek, ktoré sa obyčajne aktivujú len v jednom mieste malej experimentálnej arény. Aktivačné polia mriežkovej bunky nie sú v prostredí rozložené náhodne, ale sú usporiadané do pravidelnej hexagonálnej mriežky (obr. 4).
Tieto bunky boli objavené v roku 2005 v entorhinálnej kôre potkanov. Entorhinálna kôra je súčasťou neokortexu a neuróny v entorhinálnej kôre priamo stimulujú hipokampus, kde sa nachádzajú miestne bunky. Vlastnosti mriežkových buniek a ich funkcia sa intenzívne študujú. Pre ich priestorovú aktivitu a priame prepojenie s hipokampom sa predpokladá, že sa podieľajú na priestorovej navigácii.
Priestorovo organizovaná aktivita miestnych buniek, buniek smeru hlavy a mriežkových buniek presvedčivo dokumentuje, že tieto neuróny spracovávajú priestorové informácie a podieľajú sa na navigácii. Fascinujúce objavy v oblasti priestorovej reprezentácie v mozgu evokujú nové otázky: Ako je priestorový signál, ktorý môžeme pri týchto bunkách pozorovať, vypočítaný v nervových okruhoch mozgu? Ako presne priestorová aktivita týchto buniek napomáha navigácii? Sú tieto neuróny a mozgové štruktúry, v ktorých sa nachádzajú, úzko špecializované len na riešenie priestorových problémov, alebo sa podieľajú aj na ďalších kognitívnych funkciách? Tieto a podobné otázky sú predmetom intenzívneho výskumu a záujmu laboratórií po celom svete. Fyziológovia sledujú aktivitu neurónov a študujú ich správanie v rôznych experimentálnych situáciách. Anatómovia študujú prepojenia medzi neuronálnymi okruhmi a anatomickú organizáciu mozgových štruktúr. Teoretickí neurovedci vyvíjajú a testujú modely neurónových okruhov podieľajúcich sa na navigácii, aby lepšie porozumeli spracúvaniu priestorovej informácie. Tento výzkum je významný nielen z hľadiska porozumenia priestorovej navigácii, ale ponúka možnosť porozumieť tomu, ako sa v našich mozgoch vytvárajú a používajú abstraktné modely prostredia.
Literatura
[1] E. C. Tolman, Psychological Review 55, 189–208, 1948
[2] J. O’Keefe and J. Dostrovsky, Brain Research 34, 171– 175, 1971/1
[3] J. S. Taube, R. U. Muller and J. B. Ranck Jr., Journal of Neuroscience 10, 420–435, 1990/2
[4] T. Hafting, M. Fyhn, S. Molden, M. B. Moser, E. I. Moser, Nature 436, 801–806, 2005/7052
Ke stažení
 článek ve formátu pdf [463,5 kB]
článek ve formátu pdf [463,5 kB]
O autorovi
Eduard Kelemen
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
