










Jedový aparát plazů a fylogeneze
 Plazi odjakživa fascinovali lidstvo svým bizarním vzhledem – želvy svým krunýřem, krokodýli pancéřováním a vyzbrojenou tlamou, chameleoni hned několika vnějšími znaky, hadi a někteří ještěři svým beznohým vzhledem, a zejména svou potenciální nebezpečností. Nezřídka se plazi – zejména ti nebezpeční – stávali součástí mytologie, někteří byli dokonce uctíváni jako božstva. Jedovatost byla asi vždy nejobdivuhodnějším a nejrespektovanějším plazím fenoménem. Když nahlédneme do encyklopedií obratlovců, dostane se nám zpravidla tradiční informace, že jedový aparát plazů je přítomen u několika skupin pokročilých hadů a u jediné skupiny ještěrů – korovců. Co ale vlastně o jedovém aparátu plazů doopravdy víme? Kdo je jím skutečně obdařen a jaká je jeho evoluční historie?
Plazi odjakživa fascinovali lidstvo svým bizarním vzhledem – želvy svým krunýřem, krokodýli pancéřováním a vyzbrojenou tlamou, chameleoni hned několika vnějšími znaky, hadi a někteří ještěři svým beznohým vzhledem, a zejména svou potenciální nebezpečností. Nezřídka se plazi – zejména ti nebezpeční – stávali součástí mytologie, někteří byli dokonce uctíváni jako božstva. Jedovatost byla asi vždy nejobdivuhodnějším a nejrespektovanějším plazím fenoménem. Když nahlédneme do encyklopedií obratlovců, dostane se nám zpravidla tradiční informace, že jedový aparát plazů je přítomen u několika skupin pokročilých hadů a u jediné skupiny ještěrů – korovců. Co ale vlastně o jedovém aparátu plazů doopravdy víme? Kdo je jím skutečně obdařen a jaká je jeho evoluční historie?
Klasický přístup
Jedový aparát plazů je komplexní systém tvořený modifikovaným chrupem (denticí), žlázami produkujícími jed a vlastním jedem složeným z různých enzymů, polypeptidů a jiných toxických i netoxických makromolekul, menších sloučenin, iontů a vody.
 Dle tradičních názorů jsou za jediné ještěry s jedovým aparátem považováni korovci. Korovec jedovatý (Heloderma suspectum) a korovec mexický (H. horridum) mají jedové žlázy umístěné podél spodní čelisti a modifikované zuby s rýhami pro vedení jedu. Zuby však nejsou výrazně velikostně rozlišené, jako je tomu u hadů. Někdy se spekulovalo také o jedovatosti poněkud tajemného ještěra varanovce bornejského (Lanthanotus borneensis), příbuzného varanů a korovců známého jen z několika desítek exemplářů, ale jeho jedovatost nebyla nikdy potvrzena.
Dle tradičních názorů jsou za jediné ještěry s jedovým aparátem považováni korovci. Korovec jedovatý (Heloderma suspectum) a korovec mexický (H. horridum) mají jedové žlázy umístěné podél spodní čelisti a modifikované zuby s rýhami pro vedení jedu. Zuby však nejsou výrazně velikostně rozlišené, jako je tomu u hadů. Někdy se spekulovalo také o jedovatosti poněkud tajemného ještěra varanovce bornejského (Lanthanotus borneensis), příbuzného varanů a korovců známého jen z několika desítek exemplářů, ale jeho jedovatost nebyla nikdy potvrzena.
Jedový aparát hadů se od korovčího odlišuje umístěním jedových žláz podél horní čelisti. Podle chrupu se hadi tradičně dělí do čtyř skupin.

 Aglyfní chrup mají hadi, u kterých se nenacházejí zuby speciálně upravené k aplikování jedu čili zuby s rýhou na povrchu nebo kanálkem uvnitř. Do této kategorie náleží všichni evolučně původní hadi (od slepáků přes vinejše a krátkorepy až k různým hroznýšovitým), dále také bradavičníci a řada užovek. U mnoha z nich sice existuje diferencovaný chrup, ale pokud nejsou přítomny zmiňované rýhy nebo kanálky, stále jde o chrup aglyfní. Speciálním typem aglyfního chrupu je chrup opistodontní, u něhož jsou v zadní části horní čelisti zvětšené zuby, ale bez rýh. Takový typ je častý u řady užovek a bývá mylně zaměňován s chrupem opistoglyfním.
Aglyfní chrup mají hadi, u kterých se nenacházejí zuby speciálně upravené k aplikování jedu čili zuby s rýhou na povrchu nebo kanálkem uvnitř. Do této kategorie náleží všichni evolučně původní hadi (od slepáků přes vinejše a krátkorepy až k různým hroznýšovitým), dále také bradavičníci a řada užovek. U mnoha z nich sice existuje diferencovaný chrup, ale pokud nejsou přítomny zmiňované rýhy nebo kanálky, stále jde o chrup aglyfní. Speciálním typem aglyfního chrupu je chrup opistodontní, u něhož jsou v zadní části horní čelisti zvětšené zuby, ale bez rýh. Takový typ je častý u řady užovek a bývá mylně zaměňován s chrupem opistoglyfním.
Opistoglyfní chrup je charakteristický zvětšenými zuby s rýhou, umístěnými v zadní části horní čelisti. Vyskytuje se rovněž u mnoha druhů užovek a suchozemské formy s tímto chrupem byly dříve řazeny do společné podčeledi a všeobecně nazývány bojgy. Ve skutečnosti toto jméno přísluší jen rodu Boiga, jak si ještě vysvětlíme. Opistoglyfní chrup mají také vodnářky, sladkovodní hadi z jihovýchodní Asie (neplést s mořskými vodnáři).
 Další dva typy chrupu se vyznačují jedovými zuby s kanálkem, umístěnými v přední části úst. V jednom případě jsou tyto zuby poměrně krátké a pevně přisedlé vpředu na čelisti – typ proteroglyfní, typický pro hady korálovcovité (např. korálovce, kobry, mamby, bungary, pakobry, mořské hady – vodnáře a vlnožily). V druhém případě jsou jedové zuby dlouhé a rovněž pevně přisedlé na čelisti, ta je však extrémně zkrácená a pohyblivá a umožňuje sklopit jedové zuby do klidové polohy podél patra. Tento typ, nazývaný solenoglyfní, je typický pro hady zmijovité (zmije, chřestýše a jejich příbuzné) a pro poněkud zvláštní zemězmije (rod Atractaspis).
Další dva typy chrupu se vyznačují jedovými zuby s kanálkem, umístěnými v přední části úst. V jednom případě jsou tyto zuby poměrně krátké a pevně přisedlé vpředu na čelisti – typ proteroglyfní, typický pro hady korálovcovité (např. korálovce, kobry, mamby, bungary, pakobry, mořské hady – vodnáře a vlnožily). V druhém případě jsou jedové zuby dlouhé a rovněž pevně přisedlé na čelisti, ta je však extrémně zkrácená a pohyblivá a umožňuje sklopit jedové zuby do klidové polohy podél patra. Tento typ, nazývaný solenoglyfní, je typický pro hady zmijovité (zmije, chřestýše a jejich příbuzné) a pro poněkud zvláštní zemězmije (rod Atractaspis).
Fylogenetický pohled na (jedovaté) hady

 Dodnes přežívá mylná představa o postupné evoluci chrupu z aglyfního přes opistoglyfní a proteroglyfní až k solenoglyfnímu. Moderní doba však přináší moderní metody. V oboru třídění organizmů je už delší dobu populární fylogenetická systematika a s ní související interpretace evoluční historie morfologických či jiných fenotypových znaků. Fylogenetická systematika by nám měla v ideálním případě ukazovat obraz skutečných příbuzenských vztahů a ne jen podobností, čímž byla a stále ještě je zatížena systematika klasická, hodnotící jen znaky fenotypové. Ideální pro analýzu, i když samozřejmě ne výlučně, jsou znaky molekulárněgenetické (např. sekvence nukleotidů DNA), které nejsou tolik zatíženy adaptivní selekcí jako znaky fenotypové. 1) Jaké jsou tedy příbuzenské vztahy jedovatých i nejedovatých hadů dle výsledků nedávných fylogenetických analýz? A jak vlastně definujeme jedovaté hady?
Dodnes přežívá mylná představa o postupné evoluci chrupu z aglyfního přes opistoglyfní a proteroglyfní až k solenoglyfnímu. Moderní doba však přináší moderní metody. V oboru třídění organizmů je už delší dobu populární fylogenetická systematika a s ní související interpretace evoluční historie morfologických či jiných fenotypových znaků. Fylogenetická systematika by nám měla v ideálním případě ukazovat obraz skutečných příbuzenských vztahů a ne jen podobností, čímž byla a stále ještě je zatížena systematika klasická, hodnotící jen znaky fenotypové. Ideální pro analýzu, i když samozřejmě ne výlučně, jsou znaky molekulárněgenetické (např. sekvence nukleotidů DNA), které nejsou tolik zatíženy adaptivní selekcí jako znaky fenotypové. 1) Jaké jsou tedy příbuzenské vztahy jedovatých i nejedovatých hadů dle výsledků nedávných fylogenetických analýz? A jak vlastně definujeme jedovaté hady?
Tradičně se očekává, že jedovatý had by měl mít diferencované jedové zuby a žlázy produkující jed. Situace však není tak jasná a podíváme-li se na problém zeširoka, bylo by vhodnější považovat za signály jedovatosti schopnost produkovat toxiny a přítomnost serózních sekrečních buněk v horní čelisti. Všichni známí hadi s takovými znaky patří mezi užovkovce (Colubroidea), kteří tvoří nejodvozenější skupinu hadů (viz schéma 2 v rubrice Data a souvislosti příloha pdf), evolučně velmi úspěšnou a druhově bohatou. Užovkovci zahrnují asi 85 % všech žijících hadů a v tradičním systému bývají děleni do tří až čtyř čeledí: na užovkovité (Colubridae), korálovcovité (Elapidae), zmijovité (Viperidae) a zpravidla ještě zemězmijovité (Atractaspididae). Zemězmijovití jsou evolučně velmi zajímaví, ale záhadně a skrytě žijící hadi. Rod Atractaspis je charakteristický solenoglyfním chrupem (nicméně jiného typu než u zmijí), zatímco ostatní zástupci skupiny mají chrup opistoglyfní, nebo dokonce aglyfní. Navíc se k zemězmijům poslední dobou zařazuje i rod Homoroselaps, který má chrup proteroglyfní, kvůli němuž býval řazen mezi korálovcovité hady. Příbuznost druhů této napohled nesourodé skupiny však byla v nedávné době několikrát potvrzena molekulárněgenetickými metodami.
 Přední jedové zuby má jen asi 15 % druhů hadů. To znamená, že vedle relativně nepočetné skupiny evolučně primitivnějších hadů jsou ostatní hadi tzv. užovky, hadi s aglyfním nebo opistoglyfním chrupem. Již dříve se ale soudilo – a fylogenetické analýzy posledních let to potvrdily –, že užovky jsou skupina umělá. Některé jsou bližší korálovcům než jiným užovkám a některé možná i zmijím (viz schéma 3 v rubrice Data a souvislosti - příloha pdf).
Přední jedové zuby má jen asi 15 % druhů hadů. To znamená, že vedle relativně nepočetné skupiny evolučně primitivnějších hadů jsou ostatní hadi tzv. užovky, hadi s aglyfním nebo opistoglyfním chrupem. Již dříve se ale soudilo – a fylogenetické analýzy posledních let to potvrdily –, že užovky jsou skupina umělá. Některé jsou bližší korálovcům než jiným užovkám a některé možná i zmijím (viz schéma 3 v rubrice Data a souvislosti - příloha pdf).
Jedové žlázy užovkovců
 Přítomnost žlázy není nutně vázána na diferencovaný chrup horní čelisti a naopak. Za původní je považována sériově uspořádaná hornoretní žláza, která je přítomna u skupiny Xenodermatinae. Téměř u všech ostatních skupin „užovek“ – užovkovců s aglyfním nebo opistoglyfním chrupem – je po stranách horní čelisti přítomna Duvernoyova žláza, která produkuje toxiny. Morfologicky a fyziologicky je odlišná od pravé jedové žlázy hadů s předními jedovými zuby, předpokládá se však, že je s ní homologická. Jen u nemnoha „užovek“ Duvernoyova žláza chybí. Protože je to spíš výjimečný stav, je nejpravděpodobnějším vysvětlením sekundární ztráta. 2)
Přítomnost žlázy není nutně vázána na diferencovaný chrup horní čelisti a naopak. Za původní je považována sériově uspořádaná hornoretní žláza, která je přítomna u skupiny Xenodermatinae. Téměř u všech ostatních skupin „užovek“ – užovkovců s aglyfním nebo opistoglyfním chrupem – je po stranách horní čelisti přítomna Duvernoyova žláza, která produkuje toxiny. Morfologicky a fyziologicky je odlišná od pravé jedové žlázy hadů s předními jedovými zuby, předpokládá se však, že je s ní homologická. Jen u nemnoha „užovek“ Duvernoyova žláza chybí. Protože je to spíš výjimečný stav, je nejpravděpodobnějším vysvětlením sekundární ztráta. 2)
Pravé jedové žlázy se vyskytují jen u hadů s předními jedovými zuby (tedy u zmijovitých, korálovcovitých a zemězmijovitých) a pro každou skupinu jsou charakteristické několika specifickými znaky.
Jedový chrup užovkovců
Opistoglyfní chrup v kombinaci s Duvernoyovou žlázou nelze považovat za primitivní. Využívá ho úspěšně řada druhů několika blíže nepříbuzných skupin užovkovců a různých modifikací zvětšených horních zubů je nepřeberně. Teprve v posledních letech se ukazuje (např. díky elektronové mikroskopii), že rýhy či naopak lišty mohou být přítomny z různých stran na zubech jakékoliv velikosti a pozice na čelisti. Takto modifikované zuby lze najít u většiny podčeledí „užovek“ (v linii korálovců, pravých užovek i vodnářek), tudíž podčeleď bojg pojímaná donedávna na základě opistoglyfního chrupu naprosto nevyhovuje.
 Oba typy jedového aparátu s předními jedovými zuby mají zřejmě společný evoluční původ u dávného společného předka, který měl serózní sekreční buňky a pravděpodobně diferencovaný maxilární chrup. Nicméně proteroglyfní chrup korálovcovitých a solenoglyfní chrup zmijovitých měly následně nezávislou evoluci. Stejně tak byl nezávislý vývoj solenoglyfního systému zemězmijů a zmijovitých. Rovněž proteroglyfní uspořádání u rodu Homoroselaps mohlo vzniknout nezávisle na evoluci korálovcovitých. Přesné evoluční vztahy mezi korálovcovitými a zemězmijovitými ale zůstávají nejasné, takže nemůžeme zatím s jistotou říci, zda se systém předních jedových zubů vyvinul dvakrát, třikrát nebo dokonce čtyřikrát. Z výsledků fylogenetických analýz je také patrné, že zmijovití nejsou evolučně nejpokročilejší skupinou hadů, přestože se jejich jedový aparát považuje za nejdokonalejší. A členění hadího chrupu na obvyklé čtyři typy je z evolučního pohledu bezpředmětné.
Oba typy jedového aparátu s předními jedovými zuby mají zřejmě společný evoluční původ u dávného společného předka, který měl serózní sekreční buňky a pravděpodobně diferencovaný maxilární chrup. Nicméně proteroglyfní chrup korálovcovitých a solenoglyfní chrup zmijovitých měly následně nezávislou evoluci. Stejně tak byl nezávislý vývoj solenoglyfního systému zemězmijů a zmijovitých. Rovněž proteroglyfní uspořádání u rodu Homoroselaps mohlo vzniknout nezávisle na evoluci korálovcovitých. Přesné evoluční vztahy mezi korálovcovitými a zemězmijovitými ale zůstávají nejasné, takže nemůžeme zatím s jistotou říci, zda se systém předních jedových zubů vyvinul dvakrát, třikrát nebo dokonce čtyřikrát. Z výsledků fylogenetických analýz je také patrné, že zmijovití nejsou evolučně nejpokročilejší skupinou hadů, přestože se jejich jedový aparát považuje za nejdokonalejší. A členění hadího chrupu na obvyklé čtyři typy je z evolučního pohledu bezpředmětné.
Nejedovatá nebo jedovatá užovka?
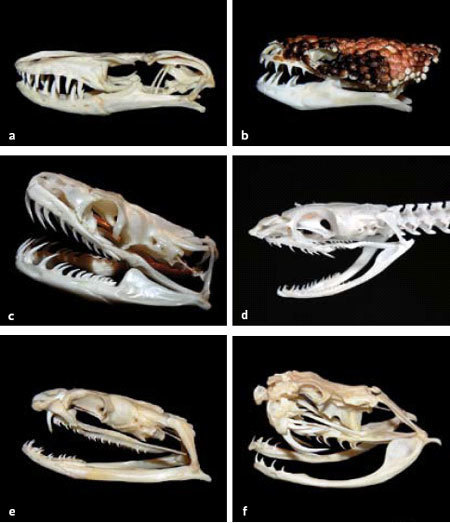 Jak jsme si již vysvětlili, pro tvorbu toxinů nejsou nezbytné jedové či Duvernoyovy žlázy, ale serózní sekreční buňky. A zdá se, že právě takové buňky, uspořádané do malých žlázek, byly dlouho přehlíženy. Přelomovým objevem byl nález neurotoxinu u pravé užovky Coelognathus radiatus. Tu běžně chovají i čeští teraristé (i když ji znají spíše pod starším jménem Elaphe radiata) a všeobecně ji považují za nejedovatou. Bryan G. Fry a jeho spolupracovníci izolovali z jejích ústních výměšků neurotoxin a zjistili, že je příbuzný rodině neurotoxinů 3FTX, považovaných za výlučný (apomorfní) znak korálovcovitých hadů. Pojmenovali ho α-colubritoxin. Jeho funkční význam je však u tohoto druhu patrně minimální. Přítomnost stejných neurotoxinů pak byla následně potvrzena také u dalších skupin užovkovců, včetně poměrně původních vodnářek. To dokazuje, že neurotoxiny 3FTX vznikly daleko dříve, již v rané evoluci užovkovců, a schopnost jejich tvorby si zachovaly i druhy s redukovaným jedovým aparátem.
Jak jsme si již vysvětlili, pro tvorbu toxinů nejsou nezbytné jedové či Duvernoyovy žlázy, ale serózní sekreční buňky. A zdá se, že právě takové buňky, uspořádané do malých žlázek, byly dlouho přehlíženy. Přelomovým objevem byl nález neurotoxinu u pravé užovky Coelognathus radiatus. Tu běžně chovají i čeští teraristé (i když ji znají spíše pod starším jménem Elaphe radiata) a všeobecně ji považují za nejedovatou. Bryan G. Fry a jeho spolupracovníci izolovali z jejích ústních výměšků neurotoxin a zjistili, že je příbuzný rodině neurotoxinů 3FTX, považovaných za výlučný (apomorfní) znak korálovcovitých hadů. Pojmenovali ho α-colubritoxin. Jeho funkční význam je však u tohoto druhu patrně minimální. Přítomnost stejných neurotoxinů pak byla následně potvrzena také u dalších skupin užovkovců, včetně poměrně původních vodnářek. To dokazuje, že neurotoxiny 3FTX vznikly daleko dříve, již v rané evoluci užovkovců, a schopnost jejich tvorby si zachovaly i druhy s redukovaným jedovým aparátem.
Raný vznik jedového aparátu aneb Jedovatí varani a jedovaté agamy
 Ještě význačnější objev však na sebe nenechal dlouho čekat. Fry se rozhodl ve svých výzkumech „nejedovatých“ plazů pokračovat a ověřit již starší hypotézu (založenou na praxi chovatelů a herpetologů) o jisté jedovatosti slin varanů, která však nebyla nikdy řádně studována. Fryův tým nejenže potvrdil produkci toxinů, ale také otestoval jejich účinky, fylogeneticky analyzoval jejich sekvenci aminokyselin, a navíc našel patřičné jedové žlázy. Jsou dokonce podobné žlázám korovců, jsou rovněž uloženy v dolní čelisti a také mají dutinu pro uskladnění jedu.
Ještě význačnější objev však na sebe nenechal dlouho čekat. Fry se rozhodl ve svých výzkumech „nejedovatých“ plazů pokračovat a ověřit již starší hypotézu (založenou na praxi chovatelů a herpetologů) o jisté jedovatosti slin varanů, která však nebyla nikdy řádně studována. Fryův tým nejenže potvrdil produkci toxinů, ale také otestoval jejich účinky, fylogeneticky analyzoval jejich sekvenci aminokyselin, a navíc našel patřičné jedové žlázy. Jsou dokonce podobné žlázám korovců, jsou rovněž uloženy v dolní čelisti a také mají dutinu pro uskladnění jedu.
 Při laboratorních testech toxinů druhu Varanus varius se podařilo prokázat významný vliv na pokles krevního tlaku a srážení krve čili bioaktivity spojené s rychlou ztrátou vědomí a krvácením. Ze záznamů pokousaných chovatelů jsou známy další symptomy – otok, vystřelující bolest, závratě, slabost svalů, ztížené dýchaní, tachykardie. Ještě překvapivější byl nález jedových žlázek u agam, konkrétně u agamy vousaté (Pogona barbata). Její drobné žlázky jsou sériově uspořádané, laločnaté, přítomné jak v dolní, tak v horní čelisti. Sekrece sice nebyla dostatečná pro laboratorní testování účinku toxinů, stačila však pro izolaci toxinů za účelem sekvenace a následné analýzy příbuzenských vztahů, které potvrdily, stejně jako v případě varanů, příbuznost s toxiny korovců i různých jedovatých hadů včetně kober, mamb a chřestýšů.
Při laboratorních testech toxinů druhu Varanus varius se podařilo prokázat významný vliv na pokles krevního tlaku a srážení krve čili bioaktivity spojené s rychlou ztrátou vědomí a krvácením. Ze záznamů pokousaných chovatelů jsou známy další symptomy – otok, vystřelující bolest, závratě, slabost svalů, ztížené dýchaní, tachykardie. Ještě překvapivější byl nález jedových žlázek u agam, konkrétně u agamy vousaté (Pogona barbata). Její drobné žlázky jsou sériově uspořádané, laločnaté, přítomné jak v dolní, tak v horní čelisti. Sekrece sice nebyla dostatečná pro laboratorní testování účinku toxinů, stačila však pro izolaci toxinů za účelem sekvenace a následné analýzy příbuzenských vztahů, které potvrdily, stejně jako v případě varanů, příbuznost s toxiny korovců i různých jedovatých hadů včetně kober, mamb a chřestýšů.
 Hlavním výstupem objevu je pak hypotéza, že se jedový aparát vyvinul asi před 200 miliony let, již v rané evoluční historii šupinatých plazů, a sice někde u společného předka skupiny Anguimorpha (např. varani) – Iguania (např. agamy) – Serpentes (hadi), pojmenované Toxicofera („nesoucí jed“). Tento objev dobře nahrál výsledkům nedávných molekulárních analýz šupinatých plazů, které právě na blízký příbuzenský vztah tří skupin ukázaly (zcela nečekaně oproti tradičnímu přístupu; viz schéma 1 v rubrice Data a souvislosti - příloha pdf). Z práce Frye a jeho kolegů bohužel není zcela jasné, zda autoři hledali jedové žlázy a toxiny také u jiných zástupců šupinatých plazů, jak z vnitřku skupiny Toxicofera, např. v rámci slepýšovitých (Anguidae), tak pro negativní kontrolu u zástupců mimo Toxicofera.
Hlavním výstupem objevu je pak hypotéza, že se jedový aparát vyvinul asi před 200 miliony let, již v rané evoluční historii šupinatých plazů, a sice někde u společného předka skupiny Anguimorpha (např. varani) – Iguania (např. agamy) – Serpentes (hadi), pojmenované Toxicofera („nesoucí jed“). Tento objev dobře nahrál výsledkům nedávných molekulárních analýz šupinatých plazů, které právě na blízký příbuzenský vztah tří skupin ukázaly (zcela nečekaně oproti tradičnímu přístupu; viz schéma 1 v rubrice Data a souvislosti - příloha pdf). Z práce Frye a jeho kolegů bohužel není zcela jasné, zda autoři hledali jedové žlázy a toxiny také u jiných zástupců šupinatých plazů, jak z vnitřku skupiny Toxicofera, např. v rámci slepýšovitých (Anguidae), tak pro negativní kontrolu u zástupců mimo Toxicofera.
Z tohoto pohledu již v budoucnu asi nepřekvapí, jestliže se přece jen potvrdí jedovatost varanovce bornejského nebo dalších a dalších druhů varanů. Rozhodně by však bylo zajímavé ověřit a prostudovat, v jakém rozsahu je přítomen jedový aparát u anguidů, včetně např. našeho slepýše křehkého (Anguis fragilis) či dalších druhů agam, leguánů a chameleonů.
Nutno ovšem poznamenat, že stále ještě existují i jiné pohledy na fylogenezi šupinatých a evoluci jedového aparátu. Jedním takovým „narušitelem“ je například hypotéza monofyletičnosti ještěrů (s dvouplazy jako vnitřní skupinou) a hadů jako jejich sesterskou skupinou, jak ukazují výsledky analýz kompletního mitochondriálního genomu. Rovněž pro morfology je asi těžko přípustná představa, že by iguánie s měkkým jazykem (podobně jako haterie) mohly být vnitřní skupinou skleroglos a nikoli bazální skupinou šupinatých, jak se dlouhá léta předpokládalo.
Literatura
Fry B. G. et al.: Isolation of a Neurotoxin (ácolubritoxin) from a Nonvenomous Colubrid: Evidence for Early Origin of Venom in Snakes, J. Mol. Evol. 57, 446–452, 2003Fry B. G. et al.: Early evolution of the venom system in lizards and snakes, Nature 439, 584–588, 2006
Jackson K.: The evolution of venomdelivery systems in snakes, Zool. J. Linn. Soc. 137, 337–354, 2003
Lawson R. et al.: Phylogeny of the Colubroidea (Serpentes): New evidence from mitochondrial and nuclear genes, Mol. Phyl. Evol. 37, 581–601, 2005
Vidal N.: Colubroid Systematics: Evidence for an early appearance of the venom apparatus followed by extensive evolutionary tinkering, J. Toxicol.Toxin Reviews 21, 21–41, 2002
Vidal N., Hedges S. B.: Higherlevel relationships of (caenophidian) snakes inferred from four nuclear and mitochondrial genes, C. R. Biologies 325, 977–985 a 987–995, 2002
Vidal N., Hedges S. B.: Molecular evidence for a terrestrial origin of snakes, Proc. R. Soc. Lond. B (Suppl.) 271, 226–229, 2004
Vidal N., Hedges S. B.: The phylogeny of squamate reptiles (lizards, snakes, and amphisbaenians) inferred from nine nuclear proteincoding genes, C. R. Biologies 328, 1000–1008, 2005
Poznámky
Mořští hadi
 Mořští hadi jsou všeobecně známí svým vysokým toxickým potenciálem. Je to dáno již tím, že jde o korálovcovité hady, kteří disponují velmi účinnými neuro toxiny typu 3FTX. Jed se v evoluci dále vyvíjel a specializoval se na paralyzaci ryb – rychlé kořisti, která může ve vodním prostředí snadno uniknout. Ještě zajímavější je však druhotná redukce až téměř ztráta jedového aparátu u druhů, které se specializovaly na požírání jiker – vodnářů Aipysurus eydouxii a Emydocephalus annulatus. Jejich jed je asi stokrát méně toxický než u příbuzných druhů živících se rybami. U druhu A. eydouxii se zjistilo, že za inaktivací toxinu stojí jediná mutace genu odpovědného za tvorbu neurotoxinu 3FTX. Jde o deleci dvou nukleotidů, která způsobila posun čtecího rámce na patřičném genu čili totální změnu a inaktivaci výsledného toxinu.
Mořští hadi jsou všeobecně známí svým vysokým toxickým potenciálem. Je to dáno již tím, že jde o korálovcovité hady, kteří disponují velmi účinnými neuro toxiny typu 3FTX. Jed se v evoluci dále vyvíjel a specializoval se na paralyzaci ryb – rychlé kořisti, která může ve vodním prostředí snadno uniknout. Ještě zajímavější je však druhotná redukce až téměř ztráta jedového aparátu u druhů, které se specializovaly na požírání jiker – vodnářů Aipysurus eydouxii a Emydocephalus annulatus. Jejich jed je asi stokrát méně toxický než u příbuzných druhů živících se rybami. U druhu A. eydouxii se zjistilo, že za inaktivací toxinu stojí jediná mutace genu odpovědného za tvorbu neurotoxinu 3FTX. Jde o deleci dvou nukleotidů, která způsobila posun čtecího rámce na patřičném genu čili totální změnu a inaktivaci výsledného toxinu.Recentní mořští hadi patří do dvou skupin – vodnáři (několik rodů) a vlnožilové (rod Laticauda). Dříve se řadili do společné čeledi vodnářovitých, dnes již ale víme, že všichni mořští hadi jsou vnitřní skupinou hadů korálovcovitých (Elapidae), konktrétně australo-papuánsko-melanéské skupiny. Vlnožilové se od vodnářů výrazně liší například tím, že samice vlnožilů vylézají na povrch klást vejce, zatímco vodnáři rodí živá mláďata přímo v moři. Dlouho se ale nevědělo, v jakém příbuzenském vztahu obě skupiny jsou a jaký přesně mají vztah k pozemním formám. Studie, která byla publikována nedávno, ukázala, že vlnožilové jsou příbuzní melanéské skupině korálovcovitých ze Šalamounových ostrovů, která se zdá být vůči všem ostatním australo-papuánským hadům (včetně vodnářů) sesterská. Australo-papuánští korálovcovití pak tvoří tři linie – velké vejcorodé druhy, malé vejcorodé druhy a živorodé druhy, kam patří také živorodí vodnáři. Vlnožilové a vodnáři si tedy skutečně nejsou blízce příbuzní. Vlnožilové vznikli již na počátku radiace korálovcovitých v australo-papuánsko-melanéské oblasti, zatímco vodnáři jsou výsledkem radiace poměrně nedávné. Zvlášť rychlá adaptivní radiace proběhla v druhově početné linii kolem nepřirozeného rodu „Hydrophis“. Zde speciace probíhala zřejmě v souladu s historickým kolísáním mořské hladiny, které způsobovalo dočasné přerušování toku genů mezi dočasně oddělenými populacemi.
Ke stažení
 článek v souboru pdf [1,03 MB]
článek v souboru pdf [1,03 MB]- příloha v souboru pdf [1,05 MB]
O autorovi
Václav Gvoždík
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
