









RNAi
Nobelovu cenu za fyziologii a lékařství v roce 2006 získali Andrew Fire a Craig Mello za objev fenoménu interference RNA (RNAi) u háďátka Caenorhabditis elegans. 1) Roku 1998 ukázali, že vnesení dvouvláknové RNA (dsRNA) do červíků způsobuje vývojové defekty, jaké bychom očekávali po odstranění genů odpovídajících svou sekvencí dotyčné dsRNA. Háďátka vypadala jako mutanti postrádající určitý gen, přestože žádný zásah do jejich DNA nebyl proveden. Brzy vyšlo najevo, že dsRNA blokuje aktivitu čili expresi genů až po jejich přepisu do formy mRNA. Spustí enzymatickou degradaci mRNA právě toho genu, jenž svou sekvencí dsRNA přesně odpovídá (obrázek 1). Jinými slovy, pomocí RNAi můžeme v živém organizmu umlčet libovolný gen pouze na základě znalosti jeho sekvence a podobně jako u mutantů nalézt podle vyvolané fenotypické změny jeho funkci. Rozdíl je v tom, že zatímco přípravou mutantních organizmů trávíme měsíce a někdy roky, dsRNA vyrobíme ve zkumavce za hodinu.
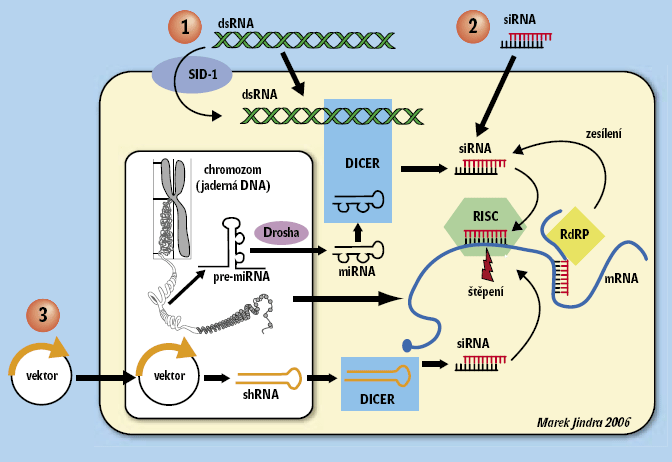 Zpráva se rozšířila rychlostí blesku, a ještě dříve, než byl mechanizmus RNAi zřejmý, začala vlna výzkumníků využívat RNAi k řešení nejrůznějších biologických otázek. Ukázalo se, že kromě háďátka funguje RNAi také u dalších organizmů včetně lidských buněk. Za osm let, po které je metoda RNAi dostupná, bylo publikováno přes jedenáct tisíc vědeckých studií, které RNAi nějakým způsobem využívají (obrázek 2).
Zpráva se rozšířila rychlostí blesku, a ještě dříve, než byl mechanizmus RNAi zřejmý, začala vlna výzkumníků využívat RNAi k řešení nejrůznějších biologických otázek. Ukázalo se, že kromě háďátka funguje RNAi také u dalších organizmů včetně lidských buněk. Za osm let, po které je metoda RNAi dostupná, bylo publikováno přes jedenáct tisíc vědeckých studií, které RNAi nějakým způsobem využívají (obrázek 2).
Cílené potlačování genové exprese bylo známo již před rokem 1998, zejména u rostlin. Například kosuprese uměle vloženou (transgenní) DNA, potlačování aktivity virů a transpozonů nebo blokování translace pomocí antisense nukleotidových sekvencí 2) patří mezi dlouho známé principy umlčování genů, jejichž mechanizmus je navíc RNAi příbuzný (pojítkem je vznik dsRNA). Jakkoli byly tyto poznatky zásadní, k Nobelově ceně nesměřovaly. Teprve práce na C. elegans totiž změnila svět. Fire a Mello měli výhodu, že jejich model rychle a přesvědčivě ukázal důležitost RNAi, přestože autoři zpočátku nevěděli, co vlastně objevili.
Ačkoli metodický přínos RNAi je obrovský, není to jen technologie. Mello sám říká, že skutečný objev přišel až s pochopením mechanizmu RNAi. Ukázalo se, že dsRNA není lidským vynálezem, ale že organizmy využívaly RNAi odpradávna např. jako obranu proti virům. Objev RNAi katalyzoval také nečekané zjištění, že buňky většiny organizmů obsahují mnoho malých molekul RNA, jejichž úkolem není produkce bílkovin, ale řízení aktivity ostatních genů procesem RNAi (obrázek 1). Zatímco před pár lety neměl o těchto mikroRNA (miRNA) nikdo ani ponětí, dnes se zdá, že v živém organizmu není děj, do něhož by nezasahovaly.
V tomto článku nelze postihnout všechny aspekty RNAi ani její terapeutický potenciál, který se už začíná projevovat například při úspěšném potlačování nádorů. Proto se soustředíme na oblast základního výzkumu, kde je přínos RNAi klíčový. Schopnost umlčovat jednotlivé geny nám totiž především ukazuje, co mají tyto geny za úkol v živém organizmu – a to je první předpoklad všech úspěšných aplikací.
Červíci a objev RNAi
 A. Fire, průkopník transformace C. elegans, pozoroval už r. 1986, že uměle vnesená DNA způsobuje u háďátek podobné změny jako mutace daného genu. Spekuloval, že z přidané DNA vzniká kromě normální (sense) mRNA také opačné vlákno (antisense), které s funkcí genu interferuje. Antisense RNA byla široce užívaným nástrojem umlčování genů – až do roku 1997. Věc měla háček: Sense RNA, která měla sloužit jako neinterferující kontrola, působila totiž stejné fenotypy jako antisense. „Nešikovní“ studenti, jejichž RNA byla více kontaminována opačným vláknem, měli nejlepší výsledky. Na jaře 1997 přijel náš kolega z konference a poradil nám: „Injikujte přímo dsRNA, je to mnohem účinnější. Jak to funguje, to se neví, ale oni na to brzo přijdou.“
A. Fire, průkopník transformace C. elegans, pozoroval už r. 1986, že uměle vnesená DNA způsobuje u háďátek podobné změny jako mutace daného genu. Spekuloval, že z přidané DNA vzniká kromě normální (sense) mRNA také opačné vlákno (antisense), které s funkcí genu interferuje. Antisense RNA byla široce užívaným nástrojem umlčování genů – až do roku 1997. Věc měla háček: Sense RNA, která měla sloužit jako neinterferující kontrola, působila totiž stejné fenotypy jako antisense. „Nešikovní“ studenti, jejichž RNA byla více kontaminována opačným vláknem, měli nejlepší výsledky. Na jaře 1997 přijel náš kolega z konference a poradil nám: „Injikujte přímo dsRNA, je to mnohem účinnější. Jak to funguje, to se neví, ale oni na to brzo přijdou.“
Přelomové články z r. 1998 ukázaly, že za domnělým artefaktem se skrývá nesmírně výkonný mechanizmus RNAi, který lze uvést do chodu poměrně dlouhými kusy dsRNA. Ty je háďátko schopno přijmout nejen po injekci do různých částí těla, ale dokonce pouhým namáčením v roztoku dsRNA nebo krmením na kultuře bakterií, které dsRNA produkují. Dvouvláknová RNA se mezi buňkami šíří, takže po jejím vpravení například do střeva můžeme pozorovat umlčení cílového genu v ostatních orgánech, a dokonce v potomstvu zasaženého červíka. Tento jev se označuje jako systemická RNAi.
„Červí komunita“ se pustila do luštění procesu RNAi pomocí genetiky. Háďátka byla zasažena mutagenem, a poté v nich byl pomocí RNAi (krmením) umlčen gen pos-1, nezbytný pro vývoj embrya. Háďátka se tak množila jedině tehdy, pokud v nich byl mutací poškozen gen, bez něhož RNAi nefunguje – jinak všechna embrya zahynula. Tak byly rychle a elegantně odhaleny geny tvořící dráhu RNAi (obrázek 1). 3)
 Systemická RNAi je velkou výhodou pro rozsáhlé studie funkce genů na úrovni celého organizmu. Několik již zveřejněných genomových analýz odhalilo například geny nezbytné pro embryogenezi, pro proces RNAi, nebo geny zúčastněné v metabolizmu tuků. Funkčně-genomické skríningy se provádějí pomocí dsRNA dodané formou krmení nebo namáčení červíků. Existují dokonce komerčně dostupné knihovny dsRNA v bakteriích, pokrývající asi 90 % genomu C. elegans.
Systemická RNAi je velkou výhodou pro rozsáhlé studie funkce genů na úrovni celého organizmu. Několik již zveřejněných genomových analýz odhalilo například geny nezbytné pro embryogenezi, pro proces RNAi, nebo geny zúčastněné v metabolizmu tuků. Funkčně-genomické skríningy se provádějí pomocí dsRNA dodané formou krmení nebo namáčení červíků. Existují dokonce komerčně dostupné knihovny dsRNA v bakteriích, pokrývající asi 90 % genomu C. elegans.
RNAi u prvoků – nečekaná variabilita
Jak již tomu v současné vědě bývá, k důležitým objevům dochází nezávisle na různých místech. V roce 1997, kdy si Fire a Mello lámali hlavu s podivným chováním RNA u háďátka, experimentovala laboratoř Elizabetty Ullu s dsRNA u jednobuněčné trypanozomy (Trypanosoma brucei), původce spavé nemoci. Po vnesení dsRNA do buněk prvoka se podařilo spustit degradaci cílové mRNA. Brzy byly vyvinuty vektory, které po přidání určité látky do kultivačního média spustí syntézu dsRNA v buňce, a tím zničí příslušnou mRNA. Dosud byly takto „vypnuty“ stovky genů trypanozomy. Výhodou RNAi není jen rychlost, ale také možnost studovat funkci proteinů nezbytných pro život. Vyřazení nepostradatelného genu mutací je pro buňku smrtelné, a žádný prvok tudíž není pro studium k dispozici. Při použití RNAi však můžeme u bičíkovce studovat fenotyp způsobený postupným ubýváním cílového proteinu až do chvíle, kdy jeho hladina klesne pod úroveň slučitelnou se životem.Fakt, že účinky RNAi u trypanozom jsou téměř totožné s účinky pozorovanými u mnohobuněčných živočichů, svědčí pro prastarý evoluční původ tohoto procesu. O to překvapivější bylo zjištění, že leishmanie (bičíkovci blízce příbuzní trypanozomě, kteří způsobují leishmaniózu) žádnou RNAi nemají, a dokonce poztrácely ze svého genomu klíčové součásti dráhy RNAi. Pro vědeckou komunitu „leishmaniologů“, vzhledem k stamilionům nemocných poměrně početnou, byl tento fakt velkým zklamáním, protože se ve funkční genomice dostali mimo hru. RNAi zřejmě chrání (snad už miliardu let) eukaryotické buňky před útokem virů a omezuje pohyb transpozonů v jejich genomu. Leishmanie proto platí za život bez RNAi vysokou aktivitou transpozonů a patrně i virů. Přesto se jim stále dobře daří. Je možné, že jim vyřazení RNAi přináší nějakou skrytou výhodu, která teprve čeká na objevení.
Vypínat geny pomocí RNAi se bohužel přes veškerou snahu zatím nedaří ani u malarických plazmodií, největších zabijáků ze společenství parazitů. Tento fakt velmi znesnadňuje studium funkcí tisíců proteinů plazmodií, které nemají žádnou podobnost s lidskými proteiny, a představují tudíž nadějný cíl pro léčiva.
Analýza genomů různých prvoků ukazuje, že základní součásti dráhy RNAi (obrázek 1) najdeme u příslušníků nejstarších evolučních větví eukaryot, např. giardií či améb, ale i rostlin. Velkým objevem současné biologie je odhalení úžasné schopnosti organizmů používat stejné geny k velmi odlišným účelům. První příklad toho, že ani komponenty dráhy RNAi nejsou výjimkou, byl nedávno popsán u nálevníků. Jak si pamatujeme ze školy, trepka má dva velmi odlišné typy jader – makronukleus a mikronukleus. Zatímco pro množení nálevníka je klíčový mikronukleus, který je podobný jádru jiných eukaryot, běžný provoz buňky obstarává makronukleus. Vědělo se, že makronukleus vzniká složitou přestavbou z mikronukleu, avšak mechanizmus přestavby zůstával hádankou. Až nedávné studie ukázaly, že jedinečně pozměněný mechanizmus RNAi je schopen odstranit obrovské množství DNA tak, aby do makronukleu přešla jen část genomu. U prvoků, kteří představují největší genetickou diverzitu v rámci eukaryot, bude pravděpodobně odhalen multifunkční charakter RNAi.
Mouchy a brouci – reverzní genetika bez hranic
Sto let genetického výzkumu udělalo z malé mušky octomilky (Drosophila melanogaster) biologický supermodel. Původním přístupem byla tradiční genetika, při níž experimentátor nejprve na základě změn fenotypu vyselektoval mutantní linie much, a teprve později (někdy o celá desetiletí) byly mutace ztotožněny s odpovědnými geny. Roku 1982 zavedli G. Rubin a A. C. Spradling transformaci, která umožnila vnášet cizí DNA do genomu mušek a později expresi nově vnesených genů také tkáňově specificky řídit. Jak uvidíme, tato schopnost přináší velkou výhodu při použití RNAi.RNAi byla na mouchách poprvé úspěšně vyzkoušena hned r. 1998. J. R. Kennerdell a R. W. Carthew získali fenokopii mutace genu frizzled tím, že injikovali dsRNA tohoto genu do embryí mouchy. Když na jaře 1999 představovali své výsledky na konferenci v Seattlu, slyšeli jsme z pléna ještě námitky typu „to přece nemůže fungovat, vždyť takový jev nemá žádný biologický podklad“. Mladý laborant Kennerdell odvětil zhruba tolik, že „jemu je vcelku jedno, že to nemůže fungovat, hlavně když to funguje“. A Gregory Hannon v letech 2000–2001 definoval součásti dráhy RNAi (jako jsou Dicer nebo protein Argonaute 2 v komplexu RISC) právě na octomilce.
Pokusy s drozofilou ukázaly první úskalí RNAi. Embrya nesměla být v době injekce starší než dvě hodiny po oplození; později se vliv dsRNA neprojevil. dsRNA totiž neprostupuje buněčnými membránami, které se na konci stadia blastodermu tvoří, a tak se nedostane k cílové mRNA. V tom se drozofila liší od háďátka, kde se dsRNA mezi buňkami šíří. Omezený vstup dsRNA do buněk zůstává hlavním (a často asi jediným) problémem aplikace RNAi na další organizmy.
Dopravit dsRNA do buněk však u drozofily žádný problém není. Stačí umístit dvě kopie sekvence cílového genu do vhodného vektoru tak, aby byly v opačné orientaci, a celý konstrukt vnést do genomu mouchy pomocí výše zmíněné transformace. Po přepisu této palindromové sekvence vznikne RNA, která se na základě své vnitřní symetrie složí v dvouřetězec vyvolávající RNAi. Syntézu dsRNA, a tedy umlčení cílového genu můžeme navíc omezit na libovolnou tkáň, což nám umožňuje zjistit funkci genu v daném orgánu a zároveň obejít případy, kdy umlčení genu v celém organizmu způsobí jeho předčasnou smrt, aniž něco vypoví o pozdějším vývoji. Neschopnost dsRNA cestovat mezi buňkami se tak nakonec ukázala být výhodou, neboť získané fenotypy lze považovat za autonomní poruchu příslušných buněk.
Jinak je tomu u malého broučka potemníka
(Tribolium castaneum), který slouží jako srovnávací model pro vývojovou biologii hmyzu. Podobně jako u háďátka se dsRNAi v těle potemníka šíří přes buněčné membrány. Navíc všechna vývojová stadia – embrya, larvy, kukly i dospělí brouci – jsou citlivá na aplikaci RNAi prostou injekcí dsRNA. Funkci genů nezbytných například pro embryogenezi tak můžeme snadno studovat díky injekcím dsRNA do broučí matky; vliv umlčení genu pak pozorujeme u jejích početných vajíček. Kombinací výhod drozofily, potemníka a dalších druhů hmyzu (např. komárů a ploštic) přístupných metodám RNAi jsme dnes schopni řešit prakticky jakoukoli otázku biologie hmyzu. To byla před objevem RNAi pustá utopie.RNAi v savčích buňkách
Ačkoli i u savců lze vyvolat RNAi in vivo (např. injekcí dsRNA do embryí, elektrošokem do orgánů nebo transgenní expresí krátkého palindromu RNA), drtivá většina pokusů probíhá v buněčných kulturách. Oproti háďátku nebo hmyzu však mají diferenciované savčí buňky nevýhodu – delší kusy dsRNA v nich vyvolávají obrannou reakci, tzv. interferonovou odpověď. Ta po vstupu například virové RNA přeprogramuje genovou expresi napadené buňky tak, aby zamezila množení viru. A tím může buňku i zabít.V roce 2001 Sayda Elbashir a kolegové z týmu Thomase Tuschla ukázali, že v savčích buňkách lze geny umlčet pomocí krátké (21–22 nukleotidů) syntetické dvouvláknové siRNA o sekvenci odpovídající mRNA cílového genu. Vnesená siRNA spouští degradaci mRNA prostřednictvím stejného buněčného mechanizmu jako dsRNA (obrázek 1), nevyvolává však nežádoucí interferonovou odpověď. Přínos této metody tkví v účinnosti, jednoduchosti a použitelnosti v různých typech buněk při poměrně nízké ceně. Umlčování genů v savčích buňkách bylo totiž dosud tvrdým oříškem, neboť obvykle spoléhalo na homologní rekombinaci DNA, 4) u běžných buněčných linií velmi vzácnou.
Kvalita siRNA určuje účinnost a specifitu štěpení cílové mRNA. Pro výběr vhodné sekvence siRNA lze kromě empirických pravidel využít různé volně dostupné algoritmy. Ty však pro jednu mRNA často navrhnou odlišné siRNA, takže nezbývá než pokusem ověřit tu správnou. Zásadní je vždy potvrdit, že změny vyvolané siRNA jsou důsledkem ztráty funkce studovaného genu, a nikoli náhodného působení siRNA na jinou mRNA nebo na fyziologii buňky. Tento efekt, zvaný „off-target“, vyloučíme použitím alespoň dvou nezávislých siRNA, zacílených na různé části jedné molekuly mRNA, a kontrolní siRNA, která nemá vliv na žádný gen v buňce. Musíme samozřejmě prokázat, že hladina cílového proteinu skutečně poklesla.
Nevýhodou syntetické siRNA je její krátká životnost, která v průběhu typického pokusu (trvajícího 2–3 dny) nedovoluje cílový protein zcela odstranit. Pro trvalou expresi siRNA byly proto vyvinuty vektory, z nichž jsou přepisovány krátké palindromy RNA. Ty spontánně tvoří dsRNA ve tvaru vlásenky, kterou poté zpracovává komplex Dicer na účinně interferující molekuly siRNA. Vektory na bázi retrovirů umožňují vložení DNA do chromozomů savčích buněk. Příslušná siRNA se tak stává stabilním produktem genomu buňky a může dlouhodobě působit na cílovou mRNA.
Žhavou novinkou jsou lentivirální vektory (lentiviry jsou retroviry schopné infikovat i nedělící se buňky), které obsahují úsek příslušné siRNA vložený do původně přirozeného genu miRNA. Díky vlastnostem miRNA je siRNA vystřižena jaderným enzymem Drosha a dále exportována do cytoplazmy, kde je naštěpena na funkční siRNA. Jelikož lentiviry infikují různé typy buněk, lze jimi zprostředkovat expresi siRNA přímo v živých organizmech, např. v myších. Na trhu se dokonce objevily knihovny lentivirálních vektorů kódující experimentálně ověřené siRNA proti přibližně osmi tisícům lidských a myších genů. Je pravděpodobné, že do dvou let budou k dispozici knihovny proti všem lidským a myším genům. Experimentátor tak již nebude muset navrhovat vlastní siRNA, ale bude si moci pouze vybrat … a zaplatit.
Úspěch vektorů umožňujících RNAi přímo v tělech pokusných zvířat podnítil zájem laboratoří i biotechnologických firem o využití RNAi v medicíně. První výsledky jsou nadějné a zdá se, že doba, kdy budeme léčit rakovinu nebo virové infekce pomocí RNAi namířené proti onkogenům nebo virovým proteinům, nemusí být daleko.
Není důvod k výmluvám
Dopad objevu RNAi na vývoj vědy byl okamžitý a zásadní. RNAi nám totiž poskytla nástroj, kterým můžeme mnohem rychleji než doposud pátrat po funkci genů, a nalézt tak dosud chybějící smysl stále se hromadících sekvencí DNA z různých organizmů. Bez kauzálních genetických důkazů by tyto sekvence zůstaly jen řadou písmen. S RNAi však také něco končí: důvod k výmluvám, že funkční analýza in vivo je příliš složitá, drahá nebo metodicky nepřístupná. Po loňské Nobelově ceně zájem o využití RNAi ve světě ještě vzroste. Měli bychom se snažit, aby Česká republika v tomto směru nezaostávala.Poznámky
SLOVNÍČEK
Dicer – RNáza III, enzym štěpící dvouvláknovou RNA na kratší úseky
Drosha – RNáza, enzym upravující prekurzorovou miRNA na aktivní formu
dsRNA – dvouvláknová RNA (ds, double-stranded)
mRNA – mediátorová neboli messenger RNA, přenáší informaci z DNA na proteiny
miRNA – mikroRNA, malé molekuly RNA, které nenesou informaci o pořadí aminokyselin v proteinu, ale slouží buňce k regulaci exprese jiných genů palindrom, palindromová sekvence – vlákno RNA nebo DNA symetrické v pořadí párujících nukleotidů. Např. úsek RNA s nukleotidy AGAAUA/UAUUCU je palindromem, který se v ose symetrie (lomítko) „přeloží“ a vznikne tak molekula dsRNA, tvořená za sebou následujícími páry bází: A–U, G–C, A–U, A–U, U–A, A–U. Pokud je palindrom přerušen úsekem nepárujících nukleotidů, vznikne dvouvláknová molekula se smyčkou na konci, tzv. vlásenková RNA (shRNA).
RNAi – interference RNA
shRNA – krátká vlásenková RNA (sh, short hairpin), viz palindrom
siRNA – malá interferující RNA (si, small interfering)
Ke stažení
 článek ve formátu pdf [458,37 kB]
článek ve formátu pdf [458,37 kB]
O autorech
Vladimír Kořínek
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
