










Počátky živočišné říše
 Živočichové patří spolu s rostlinami, houbami a velkým množstvím různých „řas“ a „prvoků“ do obrovské skupiny organizmů s velkými a složitě strukturovanými buňkami, které říkáme Eukaryota. Vznik eukaryotické buňky je nesporně nejzásadnější evoluční změnou, k níž došlo od vzniku života: týká se to především tří základních okruhů znaků, úplné změny topologie buněčných membrán, reorganizace genetického materiálu a vzniku buněčné kostry neboli cytoskeletu. Podíváme-li se na naše savčí bílkoviny a s nimi spojené geny, uvidíme, že skoro čtvrtinu (23 %) z nich najdeme úplně u všech organizmů, dalších 29 % je spojeno se vznikem eukaryontní buňky a konečně 27 % se vznikem mnohobuněčných živočichů (a jen pětina bílkovin vznikala až v průběhu evoluce metazoí). Ukazuje to na zásadní význam vzniku eukaryotické buňky, srovnatelný se vznikem mnohobuněčnosti. Není to koneckonců žádná „buňka“, ale složitý komplex různých typů organizmů, totiž nukleocytoplazmy („hostitelské buňky“) a symbiotických bakteriálních mitochondrií.
Živočichové patří spolu s rostlinami, houbami a velkým množstvím různých „řas“ a „prvoků“ do obrovské skupiny organizmů s velkými a složitě strukturovanými buňkami, které říkáme Eukaryota. Vznik eukaryotické buňky je nesporně nejzásadnější evoluční změnou, k níž došlo od vzniku života: týká se to především tří základních okruhů znaků, úplné změny topologie buněčných membrán, reorganizace genetického materiálu a vzniku buněčné kostry neboli cytoskeletu. Podíváme-li se na naše savčí bílkoviny a s nimi spojené geny, uvidíme, že skoro čtvrtinu (23 %) z nich najdeme úplně u všech organizmů, dalších 29 % je spojeno se vznikem eukaryontní buňky a konečně 27 % se vznikem mnohobuněčných živočichů (a jen pětina bílkovin vznikala až v průběhu evoluce metazoí). Ukazuje to na zásadní význam vzniku eukaryotické buňky, srovnatelný se vznikem mnohobuněčnosti. Není to koneckonců žádná „buňka“, ale složitý komplex různých typů organizmů, totiž nukleocytoplazmy („hostitelské buňky“) a symbiotických bakteriálních mitochondrií.
 Ačkoli je naše znalost fylogeneze eukaryot dosti fragmentární a jaksi neusazená (to, co dnes víme, není starší než rok dva), můžeme rozumně předpokládat, že existuje šest velkých eukaryotických „říší“ – Opisthokonta, Amoebozoa, Rhizaria, Excavata (fylogeneticky asi ze všech „říší“ nejspornější), Archaeplastida (= „Plantae“) a Chromalveolata, plus několik málo izolovaných skupin, s nimiž si dosud nevíme rady (např. Apusozoa). Mezi Amoebozoa patří měňavky a hlenky i řada „bičíkovců“ (tedy jednobuněčných eukaryot, jejichž hlavním zdrojem pohybu jsou bičíky). Rhizaria představují morfologicky i ekologicky heterogenní seskupení bičíkovců a „kořenonožců“ (tedy eukaryot pohybujících se především pomocí pseudopodií neboli „panožek“), zahrnující mimo jiné i dírkonošce a mřížovce. Také Excavata jsou z větší části bičíkovci, např. trypanozomy, krásnoočka, diplomonády a trichomonády, ale patří sem i některé skupiny bývalých kořenonožců. Archaeplastida („rostliny v širokém smyslu“) zahrnují pravé rostliny včetně zelených řas, ale i ruduchy a glaukofytní řasy. Mezi Chromalveolata patří rozmanití „prvoci“, „řasy“ a „plísně“, např. rozsivky, zlativky, chaluhy, oomycety, opalinky, kryptomonády, nálevníci, obrněnky a výtrusovci. A konečně skupina Opisthokonta zahrnuje houby, mnohobuněčné živočichy (včetně „živočišných hub“) a několik skupin „prvoků“. Protože je dávným zvykem definovat „živočichy“ především ekologicky, totiž jako heterotrofní organizmy bez buněčné stěny, obvykle se živící pohlcováním celých soust potravy (fagocytózou), je třeba srovnat klasickou osádku „živočišné říše“ se skutečnými příslušníky fylogeneticky definovaných živočichů. Naprostá většina „jednobuněčných živočichů“ čili „prvoků“ totiž nemá s živočichy nic společného, tedy kromě faktu, že jsou všichni eukaryotičtí: tradiční „prvoky“ nacházíme ve skupinách Opisthokonta, Amoebozoa, Excavata, Rhizaria i Chromalveolata, prostě skoro všude.
Ačkoli je naše znalost fylogeneze eukaryot dosti fragmentární a jaksi neusazená (to, co dnes víme, není starší než rok dva), můžeme rozumně předpokládat, že existuje šest velkých eukaryotických „říší“ – Opisthokonta, Amoebozoa, Rhizaria, Excavata (fylogeneticky asi ze všech „říší“ nejspornější), Archaeplastida (= „Plantae“) a Chromalveolata, plus několik málo izolovaných skupin, s nimiž si dosud nevíme rady (např. Apusozoa). Mezi Amoebozoa patří měňavky a hlenky i řada „bičíkovců“ (tedy jednobuněčných eukaryot, jejichž hlavním zdrojem pohybu jsou bičíky). Rhizaria představují morfologicky i ekologicky heterogenní seskupení bičíkovců a „kořenonožců“ (tedy eukaryot pohybujících se především pomocí pseudopodií neboli „panožek“), zahrnující mimo jiné i dírkonošce a mřížovce. Také Excavata jsou z větší části bičíkovci, např. trypanozomy, krásnoočka, diplomonády a trichomonády, ale patří sem i některé skupiny bývalých kořenonožců. Archaeplastida („rostliny v širokém smyslu“) zahrnují pravé rostliny včetně zelených řas, ale i ruduchy a glaukofytní řasy. Mezi Chromalveolata patří rozmanití „prvoci“, „řasy“ a „plísně“, např. rozsivky, zlativky, chaluhy, oomycety, opalinky, kryptomonády, nálevníci, obrněnky a výtrusovci. A konečně skupina Opisthokonta zahrnuje houby, mnohobuněčné živočichy (včetně „živočišných hub“) a několik skupin „prvoků“. Protože je dávným zvykem definovat „živočichy“ především ekologicky, totiž jako heterotrofní organizmy bez buněčné stěny, obvykle se živící pohlcováním celých soust potravy (fagocytózou), je třeba srovnat klasickou osádku „živočišné říše“ se skutečnými příslušníky fylogeneticky definovaných živočichů. Naprostá většina „jednobuněčných živočichů“ čili „prvoků“ totiž nemá s živočichy nic společného, tedy kromě faktu, že jsou všichni eukaryotičtí: tradiční „prvoky“ nacházíme ve skupinách Opisthokonta, Amoebozoa, Excavata, Rhizaria i Chromalveolata, prostě skoro všude.
 Příbuzenské vztahy mezi eukaryotickými „říšemi“ známe velmi povrchně, především proto, že nevíme, kde se nachází „kořen“ celého stromu – neznáme tedy fylogenetické postavení společného předka všech eukaryot. Většina molekulárních analýz podporuje umístění předka někde uvnitř „říše“ Excavata, anebo – vzácněji – mezi skupinou Opisthokonta a zbytkem eukaryot. To je ovšem založeno na analýzách genových sekvencí, které snadno mohou být neblaze ovlivněny velkými rozdíly v rychlostech molekulární evoluce mezi jednotlivými fylogenetickými liniemi – což lze samozřejmě v rámci tak obrovské a prastaré skupiny, jako jsou Eukaryota, očekávat. Existují ale i znaky založené na „molekulární anatomii“, zde především na splývání různých genů. U eukaryot známe fúzi genů pro dihydrofolátreduktázu a tymidylátsyntetázu u zástupců Rhizaria, Excavata, Plantae a Chromalveolata. Fúze genů pro karbamoyl-fosfátsyntetázu II, dihydroorotázu a aspartátkarbamoyltransferázu zase naopak spojuje Amoebozoa a Opisthokonta. Výsledkem těchto analýz je tedy hypotéza předpokládající základní štěpení eukaryot na linii původně s jedním bičíkem (Amoebozoa-Opisthokonta) a linii původně se dvěma bičíky (ostatní „říše“); problém je, že zmíněná trojgenová fúze byla nalezena i u jedné ruduchy, tedy u příslušníka skupiny Archaeplastida. To nejen zpochybňuje hypotézu o příbuzenských vztazích eukaryotických „říší“, ale bohužel i víru, že fúze genů jsou tak extrémně vzácné události, že je lze bez problémů používat pro rekonstrukci fylogeneze.
Příbuzenské vztahy mezi eukaryotickými „říšemi“ známe velmi povrchně, především proto, že nevíme, kde se nachází „kořen“ celého stromu – neznáme tedy fylogenetické postavení společného předka všech eukaryot. Většina molekulárních analýz podporuje umístění předka někde uvnitř „říše“ Excavata, anebo – vzácněji – mezi skupinou Opisthokonta a zbytkem eukaryot. To je ovšem založeno na analýzách genových sekvencí, které snadno mohou být neblaze ovlivněny velkými rozdíly v rychlostech molekulární evoluce mezi jednotlivými fylogenetickými liniemi – což lze samozřejmě v rámci tak obrovské a prastaré skupiny, jako jsou Eukaryota, očekávat. Existují ale i znaky založené na „molekulární anatomii“, zde především na splývání různých genů. U eukaryot známe fúzi genů pro dihydrofolátreduktázu a tymidylátsyntetázu u zástupců Rhizaria, Excavata, Plantae a Chromalveolata. Fúze genů pro karbamoyl-fosfátsyntetázu II, dihydroorotázu a aspartátkarbamoyltransferázu zase naopak spojuje Amoebozoa a Opisthokonta. Výsledkem těchto analýz je tedy hypotéza předpokládající základní štěpení eukaryot na linii původně s jedním bičíkem (Amoebozoa-Opisthokonta) a linii původně se dvěma bičíky (ostatní „říše“); problém je, že zmíněná trojgenová fúze byla nalezena i u jedné ruduchy, tedy u příslušníka skupiny Archaeplastida. To nejen zpochybňuje hypotézu o příbuzenských vztazích eukaryotických „říší“, ale bohužel i víru, že fúze genů jsou tak extrémně vzácné události, že je lze bez problémů používat pro rekonstrukci fylogeneze.
Nás budou nejvíc zajímat Opisthokonta, kam patří i praví, tedy mnohobuněční živočichové. Je to obrovská a rozmanitá skupina zahrnující velkou většinu všech biologických druhů na naší planetě. Patří sem samostatní i koloniální bičíkovci, různé typy améb, vláknité organizmy bez bičíkatých stadií – houby – i pohyblivé mnohobuněčné organizmy s bičíkatými jednobuněčnými spermiemi – zvířata. (Právě proto, že houby jsou živočichům velmi blízké, užívání starožitného jména „houby“ pro jednu bazální skupinu živočichů, Porifera, se stává čím dál absurdnějším a konfúznějším. Proto tu zkusme pro Porifera používat už kdysi užitý český název „houbovec“, byť s jistou skepsí – tabule se asi houbovcem nikdy utírat nebude.)
I když to nevíme jistě, Opisthokonta by mohla být bazální skupina eukaryot, snad dokonce sesterská skupina všech ostatních „říší“. Nejbližšími příbuznými opistokont jsou patrně měňavky a hlenky (Amoebozoa), ať už jde o sesterskou skupinu opistokont, nebo o skupinu, která se odštěpila hned po opistokontní větvi. Z tohoto hlediska pak stojí za úvahu, zda se tato fylogenetická blízkost neprojevuje i u primitivních mnohobuněčných živočichů. U takových skupin, jako jsou houbovci, vločkovci a žahavci, se nápadně často vyskytují měňavkovité buňky s širokými, pomalu se tvořícími pseudopodii, sloužícími k pohybu i příjmu potravy (jak pěkně pravila jedna učebnice biologie, buňka jimi svou kořist oblije), a právě to, že uvolněné buňky fungují jako měňavky, jim umožňuje se zase sejít a vytvořit nového jedince.
 Všechna jednobuněčná bičíkatá stadia opistokont mají jediný „tlačný“ bičík na zadním konci pohybující se buňky; bičík svým vlněním odvádí tekutinu od buňky, čímž tlačí její tělo vpřed. Tento typ buňky je mezi eukaryoty ojedinělý (bičíky bývají obvykle aspoň dva, a pokud je vytvořen jen jeden, bývá přední, „tažný“). Další obecnou vlastností opistokont jsou převládající mitochondrie s plochými kristami (vchlípeninami vnitřní membrány mitochondrie, zvětšujícími její aktivní povrch), zatímco u ostatních organizmů převládají kristy trubicovité. Různé skupiny opistokont sdílejí i schopnost syntetizovat bílkovinu kolagen a Opisthokonta se od ostatních eukaryot liší i specifickým způsobem biosyntézy sterolů. Ovšem hlavní podporu dostala hypotéza o fylogenetické přirozenosti skupiny Opisthokonta z molekulárních studií, ať se to týká sekvencí genů pro ribozomální RNA i různé bílkoviny, nebo změn v molekulární morfologii genů (unikátní inzerce několika aminokyselin v elongačním faktoru-1α, zvláštní typ genu pro tyrozyl-tRNA-syntetázu, který patrně kdysi, na počátku evoluce opistokont, připutoval odkudsi od archebakterií, a s normální eukaryotickou tyrozyl-tRNA-syntetázou, kterou nahradil, tedy nemá nic společního). Zatím se zdá, že bazální větvení opistokont proběhlo mezi linií směřující k houbám a linií směřující k živočichům a že všichni jejich jednobuněční zástupci patří buď k jedné, nebo k druhé linii.
Všechna jednobuněčná bičíkatá stadia opistokont mají jediný „tlačný“ bičík na zadním konci pohybující se buňky; bičík svým vlněním odvádí tekutinu od buňky, čímž tlačí její tělo vpřed. Tento typ buňky je mezi eukaryoty ojedinělý (bičíky bývají obvykle aspoň dva, a pokud je vytvořen jen jeden, bývá přední, „tažný“). Další obecnou vlastností opistokont jsou převládající mitochondrie s plochými kristami (vchlípeninami vnitřní membrány mitochondrie, zvětšujícími její aktivní povrch), zatímco u ostatních organizmů převládají kristy trubicovité. Různé skupiny opistokont sdílejí i schopnost syntetizovat bílkovinu kolagen a Opisthokonta se od ostatních eukaryot liší i specifickým způsobem biosyntézy sterolů. Ovšem hlavní podporu dostala hypotéza o fylogenetické přirozenosti skupiny Opisthokonta z molekulárních studií, ať se to týká sekvencí genů pro ribozomální RNA i různé bílkoviny, nebo změn v molekulární morfologii genů (unikátní inzerce několika aminokyselin v elongačním faktoru-1α, zvláštní typ genu pro tyrozyl-tRNA-syntetázu, který patrně kdysi, na počátku evoluce opistokont, připutoval odkudsi od archebakterií, a s normální eukaryotickou tyrozyl-tRNA-syntetázou, kterou nahradil, tedy nemá nic společního). Zatím se zdá, že bazální větvení opistokont proběhlo mezi linií směřující k houbám a linií směřující k živočichům a že všichni jejich jednobuněční zástupci patří buď k jedné, nebo k druhé linii.
Jsme tedy na samém počátku živočišné říše. Kmenová linie od společného předka opistokont ke společnému předku mnohobuněčných živočichů (Metazoa) byla jistě dlouhá, bohatě rozvětvená, se spoustou „přechodných článků“, které by nám osvětlily, jak se udělalo první zvíře, ale všechny organizmy této linie mohly prostě vymřít a nezanechat ani fosilie, ani dnešní potomstvo – tedy kromě toho jediného, který se stal předkem mnohobuněčných živočichů podle první definice. Naštěstí tomu tak není – jednobuněčné organizmy, které patří do kmenové linie živočichů, existují a postupně jich dokážeme identifikovat čím dál víc. Jsou to živočichové, byť jednobuněční.
Trubénky a příbuzní
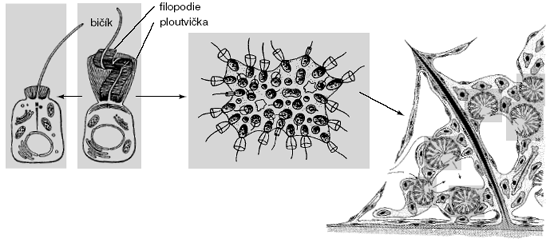
Trubénky (Choanoflagellata) jsou jednobuněčné nebo koloniální organizmy obývající mořské i sladkovodní biotopy. Jsou obvykle přisedlé, často s želatinovým obalem, někdy i s jemnými křemičitými šupinkami. Buňky v koloniích bývají propojeny cytoplazmatickými můstky, jejichž ultrastrukturu bohužel neznáme; zdá se ale, že to nejsou ta skutečná mezibuněčná spojení, jaká známe u mnohobuněčných živočichů.Nejnápadnějším znakem trubének jsou buňky s límečky: kruh 15–50 výběžků cytoplazmy, vyztužených aktinovým cytoskeletem a propojených slizovitou či vláknitou sítí, vytváří nálevku kolem dlouhého bičíku. Těmto výběžkům obvykle říkáme „mikroklky“ neboť něco podobného máme i na buňkách našeho vlastního střevního epitelu; tím ovšem dosti zatemňujeme jejich evoluční podstatu. Stejně stavěné jsou totiž tenké pseudopodie mnohých amébovitých „prvoků“, tzv. filopodie – a „mikroklky“ trubének se od jiných filopodií liší právě jenom tím, že vyrůstají v kruhu. Bičík s „ploutvičkou“ (vane) z postranních vláken mimobuněčné hmoty se od kořene ke špičce sinusoidově vlní, čímž vyhání vodu z límečku a zároveň vtahuje vodu z vnějšího prostředí mezi filopodie. Bakterie a jiné mikroskopické potravní částice se zachycují na vnějším povrchu límečku a jsou pohlcovány u báze filopodií. Kolonie trubének mohou být ploché i kulovité, s límečkovými buňkami orientovanými jak vně, tak do vnitřní dutiny, s vnitřkem kolonie vyplněným amébovými buňkami v slizovité či želatinové mimobuněčné hmotě. Taková kolonie připomíná jednoduše stavěného živočicha (povrchová vrstva buněk kolem mimobuněčné hmoty s jednotlivými buňkami), zvláště pak fragment těla houbovce. To byl také hlavní důvod, proč už staří zoologové a protozoologové předpokládali blízkou příbuznost trubének a živočichů, anebo trubének a „živočišných hub“, což bylo občas poněkud protismyslně interpretováno jako důkaz, že houbovci nejsou zvířata. K zachování monofyletických metazoí by ve skutečnosti stačilo postulovat, že ostatní mnohobuněční živočichové choanocyty druhotně ztratili. Základní morfologický typ „límečkové buňky“ (tedy bičík obklopený věncem filopodií) nacházíme v různých tkáních většiny živočišných skupin, např. i ve vylučovacích orgánech kopinatců, ale specifický tvar i potravní funkce jsou opravdu unikátní pro trubénky a houbovce. Stejně jako u metazoí i u trubének je – kolmo k bazálnímu tělísku bičíku (což je vlastně centriola) – umístěna druhá, přídatná centriola, ale nikdy tu není vytvořen příčně pruhovaný kořínek (rootlet) bičíku, který by byl srovnatelný s kořínky mnohobuněčných živočichů.
Pokud dnes víme, trubénky se množí nepohlavně, dělením buněk. U některých koloniálních trubének (Proterospongia) jsou známy složité vývojové cykly se střídáním přisedlých jednobuněčných (fáze „Choanoeca“) a mnohobuněčných (fáze „Proterospongia“), přisedlých a pohyblivých stadií. Některé přisedlé trubénky zase produkují drobné pohyblivé buňky s tlačnými bičíky, což mohou být nepohlavní spory stejně jako pohlavní buňky (gamety). Zdá se, že buňky trubének nejsou schopny dělení v bičíkatém stadiu, takže život v koloniích umožňuje počáteční specializaci na buňky pohybové a potravní, tedy s trvale přítomnými bičíky, a na dělící se, nediferencované „kmenové“ buňky uvnitř kolonie. Mimochodem – zůstalo to i nám: ani buňky mnohobuněčných živočichů nejsou schopny dělení, pokud mají bičík nebo jiný podobný výběžek s mikrotubulovou kostrou (např. axon nervových buněk).
Molekulární analýzy potvrdily, že trubénky jsou monofyletická skupina a jsou bližší živočichům než houbám. I složitě stavěná koloniální Proterospongia patří mezi pravé trubénky a rozhodně nepředstavuje žádný „přechodný článek“ mezi trubénkami a mnohobuněčnými živočichy. Spolu s trubénkami a mnohobuněčnými živočichy patří mezi Holozoa i mnohé další jednobuněčné organizmy, všechny blíže příbuzné živočichům než houbám. Dnes se to týká 4–5 skupin organizmů (všechny byly přiřazeny k trubénkám a spolu s nimi vytvořily „kmen“ Choanozoa, i když jde spíš o několik samostatných větví); mezi Opisthokonta nebo Holozoa patří zřejmě i další organizmy, které dosud málo známe, např. afelidie (Aphelidea), záhadní endoparaziti řas s opistokontními bičíkatými stadii.
Plísňovky (Mesomycetozoa = Ichthyosporea) jsou z těchto nově objevených skupin nejrozmanitější. Některé produkují spory s jedním bičíkem, u jiných bičíkatá stadia neznáme; všechny mají buněčné stěny, někdy chitinové. Mnohé druhy mají ve svém vývojovém cyklu stadia připomínající vlákna, výtrusy a cysty hub a některé také byly mezi houby tradičně řazeny, i když to byly houby podivné, bez chitinu v buněčné stěně a občas produkující amébová stadia („Enteromycetes“). Všechny druhy jsou paraziti či komenzálové obratlovců, členovců i měkkýšů. Koralochytrie (Corallochytrea) mají jemnou buněčnou stěnu a nemají bičík ani pseudopodia; jejich vývojový cyklus zahrnuje několikanásobné buněčné dělení uvnitř mateřské buňky a následné uvolňování protáhlých „spor“ skrz póry v mateřské buněčné stěně. Ministerie (Ministeriida) jsou velmi drobní a velmi podivní mořští „prvoci“ s 20 paprsčitými filopodii, která na rozdíl od trubének nevytvářejí límeček. Někdy jsou přisedlé k podkladu stvolem, jehož ultrastruktura naznačuje, že jde o modifikovaný bičík. Nukleárie (Nucleariida) jsou obvykle sladkovodní nebo půdní améby s kulovitými či zploštělými buňkami a s jemnými, paprsčitě uspořádanými pseudopodiemi. A zcela nedávno byl objeven, tentokrát v plžích, další symbiotický amébovitý organizmus, Capsaspora, původně považovaný za nukleárii, ale reprezentující další samostatnou skupinu holozoí.
Příbuzenské vztahy trubének, živočichů a všech těchto podivných „prvoků“ jsou nejasné, i když většina analýz předpokládá, že jsou bližší živočichům než houbám. Vezmeme-li v úvahu dosud spíše fragmentární molekulární i ultrastrukturální znalosti o těchto skupinách, můžeme předpokládat, že trubénky a ministerie patří skutečně na bázi mnohobuněčných živočichů (přičemž ministerie by mohly být bezprostřední sesterskou skupinou metazoí), zatímco plísňovky, kapsaspory a koralochytrie jsou umístěny bazálněji a snad tvoří monofyletickou skupinu. Ani hypotézu o monofylii skupiny Choanozoa (trubénky, ministerie, koralochytrie, plísňovky, kapsaspory) však nelze dosud považovat za zcela vyvrácenou; ovšem nukleárie jsou velmi pravděpodobně sesterskou skupinou hub, a k holozoím tedy nepatří. V každém případě je dnes těžko uniknout podezření, že i předkové holozoí byli podobnější houbám než živočichům (snad měli i buněčné stěny); ztráta „houbovitosti“ spojená s přechodem k aktivnějšímu lapání potravy pomocí filopodií mohla být prvním krokem fylogeneze trubének, ministerií a metazoí.
Trubénky jsou tedy extrémně zajímavé organizmy, zabýváme-li se vznikem mnohobuněčnosti živočichů. Proto není divu, že už začal intenzivní výzkum jejich genů a proteinů, zaměřený především na ty molekuly, jež dělají mnohobuněčné živočichy mnohobuněčnými, tedy na složitou soustavu molekul podílejících se na adhezi sousedních buněk v tkáních i na mezibuněčném transportu a mezibuněčné signalizaci. Pozoruhodné je, že značná část genů odpovědných za syntézu těchto molekul byla u trubének vskutku objevena, ačkoli jiné eukaryotické organizmy je nemají, dokonce ani tehdy, jde-li o organizmy mnohobuněčné. Tyto geny se tedy vyvinuly kdesi na bázi holozoí a pro mnohobuněčnost byly jen dodatečně použity, nikoli nutně „vynalézány“. Molekuly, které buňkám mnohobuněčných živočichů umožňují soužití s ostatními buňkami v tkáních, mohou trubénkám sloužit k získávání informací o změnách vnějšího prostředí, ke spojování buněk (při sexu, pokud ho mají, či při vzniku kolonií), či k rozpoznávání specifické kořisti. V okamžiku, kdy začala vznikat první Metazoa, byl už nezbytný repertoár mezibuněčné komunikace z velké části k dispozici.
Ke stažení
 článek ve formátu pdf [232,96 kB]
článek ve formátu pdf [232,96 kB]
O autorovi
Jan Zrzavý

Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
