









Život v půdě
 Půda v přirozených podmínkách zabezpečuje minerální výživu nejdůležitějších primárních producentů, zelených rostlin. Minerální živiny, které rostliny někdy získávají přímo zvětráváním hornin, zdaleka nedostačují jejich nárokům, neboť proces zvětrávání je sám o sobě velice pomalý. Půdní zásobu živin (horninotvorné minerály, těžko rozpustné anorganické sloučeniny, humusové látky a biogenní odumřelou hmotu) proto rostliny ještě doplňují adsorpcí značného množství aniontových a kationtových forem živin na výměnných místech půdních koloidů a nepatrným podílem živin v nesčetných sloučeninách půdního roztoku. Adsorbované živiny i živiny v půdním roztoku byly v průběhu věků uvolněny buď zvětráváním hornin, nebo v procesech rozkladu biomasy a v současnosti tvoří zásobu, jež je rostlinám více či méně rychle dostupná.
Půda v přirozených podmínkách zabezpečuje minerální výživu nejdůležitějších primárních producentů, zelených rostlin. Minerální živiny, které rostliny někdy získávají přímo zvětráváním hornin, zdaleka nedostačují jejich nárokům, neboť proces zvětrávání je sám o sobě velice pomalý. Půdní zásobu živin (horninotvorné minerály, těžko rozpustné anorganické sloučeniny, humusové látky a biogenní odumřelou hmotu) proto rostliny ještě doplňují adsorpcí značného množství aniontových a kationtových forem živin na výměnných místech půdních koloidů a nepatrným podílem živin v nesčetných sloučeninách půdního roztoku. Adsorbované živiny i živiny v půdním roztoku byly v průběhu věků uvolněny buď zvětráváním hornin, nebo v procesech rozkladu biomasy a v současnosti tvoří zásobu, jež je rostlinám více či méně rychle dostupná.
Minerální látky z mrtvých těl rostlin (nebo býložravců, kteří je zkonzumovali, popřípadě masožravců, kteří zkonzumovali ony býložravce) jsou potravou pro různé půdní organismy (naše milé „škrobáky“ a „bedrušky“ – viz předchozí díl, Vesmír 85, 212, 2006/4). Postupně se jejich činností uvolňují zpět do půdy v procesu účinné recyklace „přirozených odpadů“. Půdní mikroflóra se samozřejmě vědomě nestará o výživu rostlin, ale snaží se co nejefektivněji využít energii, která byla nahromaděna primárními producenty a kterou tyto minerální živiny v biomase „doprovázejí“.
 Podstatné je, že biomasa půdních organismů obsahuje výrazně vyšší podíl fosforu a dusíku než adekvátní biomasa rostlin. Organismy proto musí ke svému růstu získat minerální živiny navíc. Jinými slovy: na jednotku uhlíku zabudovaného do těl půdních organismů se spotřebovává více dusíku a fosforu. A protože půdních organismů je v půdě hodně a rostou rychle, minerální živiny dostupné rostlinám se mohou rychle vyčerpat. Přidáme-li do půdy třeba obilnou slámu, vytvoří se prostředí s nadbytkem uhlíku a současným nedostatkem živin. I když mikroflóra dokáže přítomné živiny účinně využívat, trpí jejich nedostatkem a přidaná sláma je rychle a neefektivně „prodýchávána“. Rostliny v takové půdě trpí nedostatkem minerální výživy až do doby, kdy je dodaná organická hmota prodýchána a systém se opět vrátí k rovnováze.
Podstatné je, že biomasa půdních organismů obsahuje výrazně vyšší podíl fosforu a dusíku než adekvátní biomasa rostlin. Organismy proto musí ke svému růstu získat minerální živiny navíc. Jinými slovy: na jednotku uhlíku zabudovaného do těl půdních organismů se spotřebovává více dusíku a fosforu. A protože půdních organismů je v půdě hodně a rostou rychle, minerální živiny dostupné rostlinám se mohou rychle vyčerpat. Přidáme-li do půdy třeba obilnou slámu, vytvoří se prostředí s nadbytkem uhlíku a současným nedostatkem živin. I když mikroflóra dokáže přítomné živiny účinně využívat, trpí jejich nedostatkem a přidaná sláma je rychle a neefektivně „prodýchávána“. Rostliny v takové půdě trpí nedostatkem minerální výživy až do doby, kdy je dodaná organická hmota prodýchána a systém se opět vrátí k rovnováze.
 „Prapůdní“ ekosystém musel existovat již v době, kdy vznikaly první rozsáhlejší porosty suchozemské vegetace. Dokladem může být minimálně devonské stáří některých půdních roztočů pancířníků (Oribatida) nebo chvostoskoků (Collembola). Půdu můžeme pravděpodobně považovat za výsledek koevoluce různých funkčních skupin organizmů v daných podmínkách. Živé organismy, přestože tvoří jen malou část půdní organické hmoty, přeměňují veškerou odumřelou organickou hmotu vstupující do půdy. Částečně ji mineralizují a částečně ji využijí v procesech biosyntézy humusových látek. V nově se tvořící půdě zpočátku převládají produkty mechanického a chemického zvětrávání. Vzniká substrát, který je odlišný od původní pevné horniny, a postupně ho osídluje mikroflóra, mezofauna a makrofauna. Připojují se k půdotvornému procesu a hrají stále důležitější roli, probíhá biologické zvětrávání. Vzniká a hromadí se humus jako standardní projev vývoje půdy. Primárně jednodušší vztahy se dolaďují až do dnešní komplexní podoby.
„Prapůdní“ ekosystém musel existovat již v době, kdy vznikaly první rozsáhlejší porosty suchozemské vegetace. Dokladem může být minimálně devonské stáří některých půdních roztočů pancířníků (Oribatida) nebo chvostoskoků (Collembola). Půdu můžeme pravděpodobně považovat za výsledek koevoluce různých funkčních skupin organizmů v daných podmínkách. Živé organismy, přestože tvoří jen malou část půdní organické hmoty, přeměňují veškerou odumřelou organickou hmotu vstupující do půdy. Částečně ji mineralizují a částečně ji využijí v procesech biosyntézy humusových látek. V nově se tvořící půdě zpočátku převládají produkty mechanického a chemického zvětrávání. Vzniká substrát, který je odlišný od původní pevné horniny, a postupně ho osídluje mikroflóra, mezofauna a makrofauna. Připojují se k půdotvornému procesu a hrají stále důležitější roli, probíhá biologické zvětrávání. Vzniká a hromadí se humus jako standardní projev vývoje půdy. Primárně jednodušší vztahy se dolaďují až do dnešní komplexní podoby.
Přestože je podíl humusových látek v půdě výrazně vyšší než podíl odumřelé i živé biomasy organismů, na celkové průměrné rychlosti obratu organického uhlíku se humusové látky podílejí méně. Důvodem je jejich podstatně pomalejší rozklad. Za předpokladu normální funkce půdního ekosystému je proto prakticky možné obsah humusových látek pokládat v krátkodobém měřítku za stálou veličinu (viz rubriku Data a souvislosti v minulém čísle, Vesmír 85, 214, 2006/4). Pro vlastní půdní metabolismus a přímou výživu rostlin mají tedy živé organismy bezprostředně vyšší význam než humusové frakce půdního uhlíku.
 Tato jednoduchá situace se však za určitých okolností může změnit. Z praxe jsou známy příklady, kdy intenzivní obhospodařování zemědělské půdy spojené s maximálním odsunem vyprodukované organické hmoty a jejím nahrazováním anorganickými hnojivy zrychlilo rozklad humusových látek. Mechanismus tohoto účinku zjevně souvisí se změnami v rovnováze živin, v provzdušnění a v mnohostranném rozrušení původní půdní struktury. Složení i aktivita živých organizmů se změní. Bude se dařit těm, kteří z půdních zásob dokážou lépe využívat právě organickou hmotu s delší dobou obratu. Je to vlastně opačný jev, než jaký jsme popisovali při tvorbě půdy: Pokud do půdy vneseme relativní nadbytek minerálních živin a strukturně ji změníme (zde orbou), půdní mikroflóra bude účinně využívat veškerou energii uloženou v organických látkách – včetně do té doby velmi těžko rozložitelných látek humusových.
Tato jednoduchá situace se však za určitých okolností může změnit. Z praxe jsou známy příklady, kdy intenzivní obhospodařování zemědělské půdy spojené s maximálním odsunem vyprodukované organické hmoty a jejím nahrazováním anorganickými hnojivy zrychlilo rozklad humusových látek. Mechanismus tohoto účinku zjevně souvisí se změnami v rovnováze živin, v provzdušnění a v mnohostranném rozrušení původní půdní struktury. Složení i aktivita živých organizmů se změní. Bude se dařit těm, kteří z půdních zásob dokážou lépe využívat právě organickou hmotu s delší dobou obratu. Je to vlastně opačný jev, než jaký jsme popisovali při tvorbě půdy: Pokud do půdy vneseme relativní nadbytek minerálních živin a strukturně ji změníme (zde orbou), půdní mikroflóra bude účinně využívat veškerou energii uloženou v organických látkách – včetně do té doby velmi těžko rozložitelných látek humusových.
Kanály a půda jako ekosystém?
 Klíčovým místem fungování půdního ekosystému je zásobník (pool) neživé organické hmoty. A klíčovým důvodem fungování procesů humifikace a mineralizace, procesů vycházejících z dekompozice této odumřelé organické hmoty, je nepředstavitelně různorodá metabolická aktivita živých (jednobuněčných, v úžasných koloniích žijících, a mnohobuněčných, krásná pletiva a tkáně vytvářejících) organismů. Z pohledu tohoto zásobníku je možno okamžitě konstatovat, že přímo do něj půdní organismy přispívají svými exkrementy (tzv. koprogenní humifikace), a stejně tak se jejich vlastní těla po své smrti stanou jeho přímou součástí. Z půdní mrtvé organické hmoty, tohoto bohatého zdroje energie a živin, se snaží čerpat velká většina půdní mikroflóry – heterotrofní půdní bakterie, aktinomycety a houby. Jejich biochemické schopnosti jsou ve srovnání s vyššími organismy naprosto fantastické. Půdní bakterie využívají nejrozmanitější formy půdní organické hmoty. Vytvářejí i společenstva, v nichž biochemické schopnosti jednotlivých druhů na sebe funkčně navazují. Role bakterií může být poněkud potlačena abiotickými faktory (klasickými příklady je snížení vlhkosti či pH), a v tom případě se začínají více uplatňovat houby.
Klíčovým místem fungování půdního ekosystému je zásobník (pool) neživé organické hmoty. A klíčovým důvodem fungování procesů humifikace a mineralizace, procesů vycházejících z dekompozice této odumřelé organické hmoty, je nepředstavitelně různorodá metabolická aktivita živých (jednobuněčných, v úžasných koloniích žijících, a mnohobuněčných, krásná pletiva a tkáně vytvářejících) organismů. Z pohledu tohoto zásobníku je možno okamžitě konstatovat, že přímo do něj půdní organismy přispívají svými exkrementy (tzv. koprogenní humifikace), a stejně tak se jejich vlastní těla po své smrti stanou jeho přímou součástí. Z půdní mrtvé organické hmoty, tohoto bohatého zdroje energie a živin, se snaží čerpat velká většina půdní mikroflóry – heterotrofní půdní bakterie, aktinomycety a houby. Jejich biochemické schopnosti jsou ve srovnání s vyššími organismy naprosto fantastické. Půdní bakterie využívají nejrozmanitější formy půdní organické hmoty. Vytvářejí i společenstva, v nichž biochemické schopnosti jednotlivých druhů na sebe funkčně navazují. Role bakterií může být poněkud potlačena abiotickými faktory (klasickými příklady je snížení vlhkosti či pH), a v tom případě se začínají více uplatňovat houby.
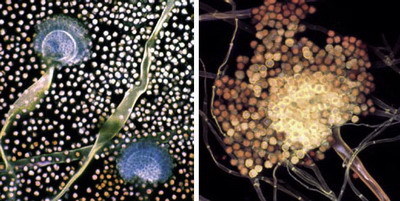 Bakteriální a houbový kanál, kterými energie a uhlíkaté látky v půdě plynou, tvoří dvě klíčové cesty rozkladu organické hmoty. O tom, zda se bude více uplatňovat jeden, nebo druhý z kanálů, rozhodují často „maličkosti“ – třeba to, jestli je půdní profil mechanicky rozrušován či nikoliv. Návaznost dalších skupin organismů na oba hlavní kanály je různá, takže poměrně nevinně vyhlížející změna obdělávání může ve svých důsledcích znamenat naprosto rozdílný vývoj daného půdního ekosystému a může vést například ke ztrátám, nebo naopak k hromadění organických látek v půdě.
Bakteriální a houbový kanál, kterými energie a uhlíkaté látky v půdě plynou, tvoří dvě klíčové cesty rozkladu organické hmoty. O tom, zda se bude více uplatňovat jeden, nebo druhý z kanálů, rozhodují často „maličkosti“ – třeba to, jestli je půdní profil mechanicky rozrušován či nikoliv. Návaznost dalších skupin organismů na oba hlavní kanály je různá, takže poměrně nevinně vyhlížející změna obdělávání může ve svých důsledcích znamenat naprosto rozdílný vývoj daného půdního ekosystému a může vést například ke ztrátám, nebo naopak k hromadění organických látek v půdě.
Na rozdíl od nadzemních ekosystémů je mnohdy téměř nemožné oddělit od sebe půdní bakterie, respektive houby a jejich potravu (mrtvou organickou hmotu neboli detritus). Většina půdních mykofágů a bakteriofágů požírá současně i značný objem detritu. V literatuře se užívá připodobnění k lidské stravě: k malému množství mikroflórového džemu musí půdní živočichové povinně zkonzumovat velké množství nechutných detritových sušenek. Striktní potravní specializace je u mnoha půdních živočichů pouze zažitou iluzí. Přesnější je antropomorfní představa, že jednotlivé druhy mají svá „oblíbená jídla“, ale když je hlad, tak snědí, co je právě po ruce. Pro funkci půdního ekosystému to má pochopitelně velký význam, neboť rychlost a způsob konzumace určité skupiny půdních mikroorganismů živočichy jsou do značné míry nezávislé na tom, jaké druhy pasoucích se živočichů jsou právě přítomny.
 V tomto bodě docházíme k zdánlivému paradoxu: půdní fauna požírá mikroflóru, a navíc jí „ujídá“ z mrtvé organické hmoty. Je jejím konzumentem a zároveň v jistém smyslu i nezvaným spolustolovníkem. To má však v mnoha případech pozitivní vliv na společenstvo půdních mikroorganismů! Způsobují to tři základní, společně působící skutečnosti:
V tomto bodě docházíme k zdánlivému paradoxu: půdní fauna požírá mikroflóru, a navíc jí „ujídá“ z mrtvé organické hmoty. Je jejím konzumentem a zároveň v jistém smyslu i nezvaným spolustolovníkem. To má však v mnoha případech pozitivní vliv na společenstvo půdních mikroorganismů! Způsobují to tři základní, společně působící skutečnosti:
– Půdní fauna udržuje mikroflóru v optimální růstové fázi, což dovoluje rychlou reakci mikroflóry na případný větší objem dostupné potravy.
– Bakterie a houby neumějí dostatečně rychle rozdrobovat mrtvé tkáně a pletiva a mohou „nasednout“ jen na omezenou plochu. Jejich růst je limitován. Půdní fauna při konzumaci mikroflóry rozdrobuje zbytky organické hmoty, a tím až o několik řádů zvětšuje povrch přístupný mikroflóře.
– Půdní živočichové mrtvou organickou hmotu při konzumaci mikrobiálně naočkují (infikují) mikroflórou – ať už z okolí či z vlastního zažívacího traktu.
Stop extrémům
Aby mohl systém dlouhodobě fungovat, potřebuje účinné regulační vazby. Zvýšené množství mrtvé organické hmoty doprovází nárůst početnosti mikroflóry. To je příklad pozitivní zpětné vazby (typu čím více, tím více), která urychluje růst populace. Kdyby působila jen sama o sobě, nastal by po vyčerpání zdrojů rozvrat společenstva. V reálném světě se však uplatňují i negativní zpětné vazby (typu čím více, tím méně), které jsou typickým atributem pastevně-kořistnických potravních řetězců, v našem případě začínajících na „loukách“ s půdní mikroflórou (viz rubriku Data a souvislosti v minulém čísle, Vesmír 85, 214, 2006/4).
 Zjednodušeně řečeno, čím více se rozvíjí mikroflóra, tím více narůstají společenstva půdní fauny (která mj. mikroflóru požírají) a tím více narůstá i počet predátorů. Několik potravních úrovní zajišťuje dostatečně účinnou regulaci. Bez ní by přidání mrtvé organické hmoty vedlo k prudkému rozvoji mikroflóry a rychlému využití hmoty, po němž by následoval prudký pokles jejího množství a kolaps půdní bioty na minimální stav. Autolyzáty a metabolity živé bioty by se do půdního prostředí uvolňovaly v obrovitých nárazech a půdní ekosystém by byl vystaven tlaku zevnitř generovaných extrémů. Půda je ale naopak prostředím překvapivě stabilním (samozřejmě z hlediska biosféry, nikoliv z hlediska např. jednotlivé kolonie nitrifikačních bakterií říčních naplavenin). Několikaúrovňová negativní zpětná vazba umožňuje v určitém rozmezí podmínek nastavit optimální rychlost zpracování mrtvé organické hmoty. Přidáním organické hmoty sice narůstá početnost mikroflóry i konzumentů, ale nijak dramaticky. V důsledku toho je substrát zpracováván pomaleji a rovnoměrněji a společenstvo, byť není v čase zcela stabilní a neměnné, má určitou dynamiku a nekolabuje.
Zjednodušeně řečeno, čím více se rozvíjí mikroflóra, tím více narůstají společenstva půdní fauny (která mj. mikroflóru požírají) a tím více narůstá i počet predátorů. Několik potravních úrovní zajišťuje dostatečně účinnou regulaci. Bez ní by přidání mrtvé organické hmoty vedlo k prudkému rozvoji mikroflóry a rychlému využití hmoty, po němž by následoval prudký pokles jejího množství a kolaps půdní bioty na minimální stav. Autolyzáty a metabolity živé bioty by se do půdního prostředí uvolňovaly v obrovitých nárazech a půdní ekosystém by byl vystaven tlaku zevnitř generovaných extrémů. Půda je ale naopak prostředím překvapivě stabilním (samozřejmě z hlediska biosféry, nikoliv z hlediska např. jednotlivé kolonie nitrifikačních bakterií říčních naplavenin). Několikaúrovňová negativní zpětná vazba umožňuje v určitém rozmezí podmínek nastavit optimální rychlost zpracování mrtvé organické hmoty. Přidáním organické hmoty sice narůstá početnost mikroflóry i konzumentů, ale nijak dramaticky. V důsledku toho je substrát zpracováván pomaleji a rovnoměrněji a společenstvo, byť není v čase zcela stabilní a neměnné, má určitou dynamiku a nekolabuje.
 Kam tím míříme? Zajímají nás dvě otázky: Co se stane, když se zastaví růst nadzemní biomasy daného stanoviště? Jak se tyto procesy promítnou do hodnocení skutečné biologické aktivity dané půdy daného stanoviště v daném čase?
Kam tím míříme? Zajímají nás dvě otázky: Co se stane, když se zastaví růst nadzemní biomasy daného stanoviště? Jak se tyto procesy promítnou do hodnocení skutečné biologické aktivity dané půdy daného stanoviště v daném čase?
Za podmínek nedostatku organické hmoty a látek vznikajících v prvních fázích jejího rozkladu se pochopitelně změní všechny původní potravní vazby typické pro přirozený půdní ekosystém. Půdní mikroflóra zpomaluje svůj růst, neboť ubývá zdroj energie. Klesá mikrobiální aktivita i početnost mikroorganizmů, a posléze i půdní fauny, neboť i té chybí dostatek potravy. Složení společenstev se výrazně mění a dominance několika málo druhů se prohlubuje. Klesá složitost vzájemných vazeb, některé vnitřní vazby půdního ekosystému se rozpadají. Snižuje se schopnost půdy tlumit vnitřní změny; to znamená, že se zvětší výkyvy podmínek půdního prostředí, vlhkosti a salinity (koncentrace půdního roztoku). A co k této neblahé situaci řekne tradiční hodnocení biologické aktivity půdy? Při povrchním sledování (např. měřením celkové produkce CO2) nemusí být tyto změny ve fungování půdního ekosystému vůbec zaznamenány. Mikroflóra v nepřítomnosti rychle dostupného půdního uhlíku intenzivněji dýchá. Půda se navenek bude chovat zdravě, i když je ve skutečnosti „ohrožena“: místo stabilizace se destabilizuje. Obsah humusových látek se snižuje, půdní struktura se ničí. Souhrnně řečeno: klesá komplexnost půdního ekosystému, která je nejlepší zárukou dlouhodobé samoregulační funkce. K tomuto tvrzení jen podotýkáme, že jsme si vědomi
– například ze studia polárních ekosystémů
– i skutečnosti, že některé ekosystémy jsou dlouhodobě stabilní, aniž by byly jejich vazby skutečně tak složité, jak popisujeme na příkladu fungování půdního ekosystému.
Prach jsi a v prach se obrátíš
 Najít několik málo, či dokonce jediný konkrétní parametr pro charakteristiku biologické aktivity půdy je naivní přání. S prohlubováním znalostí o půdě lze v úvahách dospět až do situace, kdy půdě začneme přisuzovat vlastní metabolismus (tradiční atribut živých organismů). Je nezbytné opustit snahu o stručný, jednoduchý a jednoznačný popis chování půdy. Obsah pojmu biologická aktivita by měl respektovat složitost půdních procesů a jevů. Nejlépe pracující systém je takový, který má nejlepší regulační schopnost (účinné vnitřní vazby) a v případě řady biologických systémů je zřejmé, že čím složitější systém je, tím spolehlivější je jeho autoregulace. Tak dojdeme k závěru, že vyjádření biologické aktivity by mělo nějakým způsobem zahrnovat a kvantifikovat „spolupráci“ jednotlivých složek systému. Zde může jako indikátor stavu půdy sloužit např. druhová diverzita společenstva půdních organismů.
Najít několik málo, či dokonce jediný konkrétní parametr pro charakteristiku biologické aktivity půdy je naivní přání. S prohlubováním znalostí o půdě lze v úvahách dospět až do situace, kdy půdě začneme přisuzovat vlastní metabolismus (tradiční atribut živých organismů). Je nezbytné opustit snahu o stručný, jednoduchý a jednoznačný popis chování půdy. Obsah pojmu biologická aktivita by měl respektovat složitost půdních procesů a jevů. Nejlépe pracující systém je takový, který má nejlepší regulační schopnost (účinné vnitřní vazby) a v případě řady biologických systémů je zřejmé, že čím složitější systém je, tím spolehlivější je jeho autoregulace. Tak dojdeme k závěru, že vyjádření biologické aktivity by mělo nějakým způsobem zahrnovat a kvantifikovat „spolupráci“ jednotlivých složek systému. Zde může jako indikátor stavu půdy sloužit např. druhová diverzita společenstva půdních organismů.
 Naše znalosti o toku energie jednotlivými články společenstva půdních organismů a pravidel, kterými se tento tok řídí, jsou dosud velmi nedostatečné. Je však jisté, že půda je významným úložištěm energie vázané na uhlík organických látek. Je největším zdrojem a zásobárnou uhlíku v suchozemském ekosystému (zanedbáme-li samozřejmě karbonátové horniny jako takové). Množství organického uhlíku v půdě je vyšší než celkové množství uhlíku ve vzdušném oxidu uhličitém, který se v globálním měřítku významně podílí na skleníkovém efektu. Protože je takto uskladněný uhlík z velké části pod „správou“ půdních organismů, mělo by nás zajímat jejich chování. A to i s vědomím, že naše snaha dosáhnout pravdy má stejný průběh jako snaha urychlit hmotnou částici až na rychlost světla: čím blíže jsme pravdě, tím obtížnější je postup…
Naše znalosti o toku energie jednotlivými články společenstva půdních organismů a pravidel, kterými se tento tok řídí, jsou dosud velmi nedostatečné. Je však jisté, že půda je významným úložištěm energie vázané na uhlík organických látek. Je největším zdrojem a zásobárnou uhlíku v suchozemském ekosystému (zanedbáme-li samozřejmě karbonátové horniny jako takové). Množství organického uhlíku v půdě je vyšší než celkové množství uhlíku ve vzdušném oxidu uhličitém, který se v globálním měřítku významně podílí na skleníkovém efektu. Protože je takto uskladněný uhlík z velké části pod „správou“ půdních organismů, mělo by nás zajímat jejich chování. A to i s vědomím, že naše snaha dosáhnout pravdy má stejný průběh jako snaha urychlit hmotnou částici až na rychlost světla: čím blíže jsme pravdě, tím obtížnější je postup…
HUMUS
Z jednoho úhlu pohledu jsou humusové látky jednou z nejrozšířenějších forem výskytu organického uhlíku v biosféře. Z úhlu druhého jsou jednou z nejdůležitějších půdních složek ovlivňujících půdní metabolismus (dáme-li tomuto spojení právo na život) i půdní ekosystém jako celek. Jejich tvorba začíná rozkladnými procesy organických zbytků a končí navazujícími syntézami. Rozkládány jsou v prvé řadě celulóza, hemicelulóza, lignin a proteiny, v řadě druhé pak fenolové deriváty a pryskyřice, alkaloidy, pigmenty a jiná organická barviva, taniny, tuky, vosky, oleje, keratiny, chitin, volné aminokyseliny a cukry. V podmínkách chladnějšího klimatu se během půldruhého roku rozloží některé cukry, aminokyseliny, 80–90 % proteinů a jen několik procent zrašeliněné hmoty či melaninů. Během jediného roku ztratí drtivá většina organických látek svou původní morfologickou a chemickou integritu a stává se součástí pokladnice zvané humus. Polyfenolické produkty rozkladu dále podléhají mikrobiálním oxidačním polymerizacím, při nichž vznikají makromolekuly bez vedlejších produktů, a mikrobiálním polykondenzacím, při nichž naopak dochází k odštěpení jednoduchých nízkomolekulárních produktů. Průběh obou reakcí je urychlován enzymy a některými anorganickými ionty. Produkty syntéz pak mají tendenci vytvářet velmi komplikované organické struktury, které jsou nesrovnatelně stabilnější než výchozí látky.
Různé typy humusu se ve své celkové struktuře příliš neliší. Dělí je uspořádání a četnost výskytu jejich charakteristických stavebních skupin. Takto lze humus charakterizovat jako skupinu látek na bázi rozvětvených polymerů spirální struktury s různě obsazenými funkčními skupinami. Tato představa vyhovuje i ověřenému vysokému adsorpčnímu potenciálu humusových látek. Vnější tvar těchto polymerů je flexibilní, takže i když se polymerní spirály po adsorpci na jílové minerály svinou, jejich vnitřní prostor zůstává nadále adsorpčně přístupný. Specifický povrch (vnější a vnitřní dohromady) humusových látek tak i v případě vzniku komplikovaných organominerálních komplexů zůstává obrovský.
Humus tak můžeme považovat za skupinu půdních organických sloučenin s proměnnými vnějšími rozměry (vlivem svinování a natahování polymerních řetězců), vysokým vnitřním aktivním povrchem (vlivem jeho spirální struktury) a s převažujícím negativním nábojem, a tedy i se silnou tendencí vytvářet komplexy s půdními kationty. Polymerizace (či kondenzace) a depolymerizace přitom v humusových látkách probíhají kontinuálně.
Ke stažení
 článek ve formátu pdf [1,67 MB]
článek ve formátu pdf [1,67 MB]
O autorech
Hana Šantrůčková
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
