









Lasturnatky
 Vypadají jako maličcí mlži. Dvě droboučké lasturky zpevněné svěracími svaly, sotva milimetr nebo dva nadél. Jen z úzké štěrbiny vykukují podivné článkované nožky, které tvorečka usvědčují z příslušnosti k úplně jiné živočišné skupině. Řeč je o lasturnatkách (Ostracoda), nenápadné, ale pozoruhodné skupině korýšů.
Vypadají jako maličcí mlži. Dvě droboučké lasturky zpevněné svěracími svaly, sotva milimetr nebo dva nadél. Jen z úzké štěrbiny vykukují podivné článkované nožky, které tvorečka usvědčují z příslušnosti k úplně jiné živočišné skupině. Řeč je o lasturnatkách (Ostracoda), nenápadné, ale pozoruhodné skupině korýšů.
 Lasturnatky jsou ekologicky nesmírně přizpůsobivou skupinou, která obývá téměř všechny vodní biotopy. Více než 5600 druhů žije v současnosti ve sladkých i slaných vodách, ve vlhkém prostředí afrických pralesů nebo na pobřeží Velké Británie mezi liniemi přílivu a odlivu. V České republice bylo zatím zaznamenáno 82 druhů a mnohé další mořské i sladkovodní lasturnatky lze samozřejmě nalézt ve fosilizované podobě v sedimentech od paleozoika až po kvartér.
Lasturnatky jsou ekologicky nesmírně přizpůsobivou skupinou, která obývá téměř všechny vodní biotopy. Více než 5600 druhů žije v současnosti ve sladkých i slaných vodách, ve vlhkém prostředí afrických pralesů nebo na pobřeží Velké Británie mezi liniemi přílivu a odlivu. V České republice bylo zatím zaznamenáno 82 druhů a mnohé další mořské i sladkovodní lasturnatky lze samozřejmě nalézt ve fosilizované podobě v sedimentech od paleozoika až po kvartér.
Jak to lasturnatky dělají (aby jich bylo víc)
U dnešních sladkovodních lasturnatek existují tři různé způsoby rozmnožování – bisexuální, partenogenetické a smíšené. U bisexuálních druhů jsou obě pohlaví v různém poměru přítomna v celém areálu druhu a obě se podílejí na reprodukci. Partenogenetické druhy mají čistě samičí populace. Mohou to být jednak druhy s blízkými bisexuálními příbuznými, od nichž se partenogenní druhy v minulosti pravděpodobně odštěpily, a dále druhy skupiny Darwinulocopina, které kandidují na označení dávnověcí asexuálové. 1) V současné době na světě žije již jen pozůstatek (28 druhů s 5 rody) této skupiny, velmi úspěšné a diverzifikované v mezozoiku. Na území České republiky se vyskytuje jediný recentní druh Darwinula stevensoni, v Německu v kvartérních sedimentech a recentně v Rumunsku ještě druh Microdarwinula stevensoni. V Irsku, Skotsku a Francii byl dále nalezen jinak jihoafrický a brazilský druh Penthesilenula brasiliensis. V poslední době byly díky rostoucímu zájmu o výzkum lasturnatek v Evropě nalezeny další fosilní i recentní druhy.
Mohou to být jednak druhy s blízkými bisexuálními příbuznými, od nichž se partenogenní druhy v minulosti pravděpodobně odštěpily, a dále druhy skupiny Darwinulocopina, které kandidují na označení dávnověcí asexuálové. 1) V současné době na světě žije již jen pozůstatek (28 druhů s 5 rody) této skupiny, velmi úspěšné a diverzifikované v mezozoiku. Na území České republiky se vyskytuje jediný recentní druh Darwinula stevensoni, v Německu v kvartérních sedimentech a recentně v Rumunsku ještě druh Microdarwinula stevensoni. V Irsku, Skotsku a Francii byl dále nalezen jinak jihoafrický a brazilský druh Penthesilenula brasiliensis. V poslední době byly díky rostoucímu zájmu o výzkum lasturnatek v Evropě nalezeny další fosilní i recentní druhy.
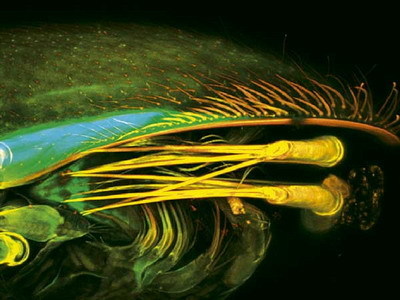 Třetím způsobem reprodukce sladkovodních lasturnatek je reprodukce smíšená. V části areálu se vyskytují samci i samice množící se bisexuálně, v jiné části pouze partenogenetické samice a ještě v jiné žijí oba reprodukční typy pospolu.
Třetím způsobem reprodukce sladkovodních lasturnatek je reprodukce smíšená. V části areálu se vyskytují samci i samice množící se bisexuálně, v jiné části pouze partenogenetické samice a ještě v jiné žijí oba reprodukční typy pospolu.  V posledním případě bisexuální populace produkují diploidní potomstvo, partenogenetické samice rovněž, 2) a krom toho se kříží se samci bisexuálních populací za vzniku triploidního potomstva samičího pohlaví. Navíc mohou v rámci smíšené reprodukcevznikat i partenogenetické populace z bisexuálních.
V posledním případě bisexuální populace produkují diploidní potomstvo, partenogenetické samice rovněž, 2) a krom toho se kříží se samci bisexuálních populací za vzniku triploidního potomstva samičího pohlaví. Navíc mohou v rámci smíšené reprodukcevznikat i partenogenetické populace z bisexuálních.
Jako smíšená reprodukce je označována rovněž geografická partenogeneze, 3) při níž čistě samičí populace převládají na většině území a bisexuální populace jsou omezeny na malé ostrůvky, většinou na jihu Evropy. Dříve se vysvětlovala rozdílnými klimatickými podmínkami v rámci areálu, např. nižšími průměrnými teplotami v severnějších oblastech. Od podobných konceptů se však upustilo – zejména s ohledem na paleontologický záznam. Ten poskytuje cenné detailní informace o historickém výskytu jednotlivých druhů s konkrétními způsoby rozmnožování. Z poměrně široké škály hypotéz vysvětlujících geografickou partenogenezi si zde stručně přiblížíme dvě nejpravděpodobnější a zároveň nejdiskutovanější – historickou a ekologickou.
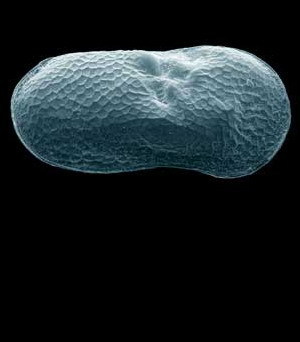 Dle historického scénáře je současné rozšíření různých populací výsledkem vývoje po poslední době ledové. Lasturnatky nemohou žít pod ledovcem nebo ve věčně zmrzlé půdě, a v glaciálu proto ustoupily před zaledněním na jih. Od počátku holocénu pak postupně znovu osídlovaly severnější oblasti. Nová území osídlují efektivněji partenogenetické samice, neboť mohou založit novou populaci z pověstného jediného vajíčka, čili jediná samice může dát vznik nové populaci. Partenogenetické populace mohou také vznikat kontinuálně z bisexuálních linií, zatímco opačný proces, vznik bisexuálních populací z partenogenetických, nebyl nikdy doložen a ani se nepředpokládá. Ekologický scénář situaci doplňuje o skutečnost, že bisexuální populace nebyly a dosud nejsou schopny v rámci areálů kompetitivně nahradit populace partenogenetické.
Dle historického scénáře je současné rozšíření různých populací výsledkem vývoje po poslední době ledové. Lasturnatky nemohou žít pod ledovcem nebo ve věčně zmrzlé půdě, a v glaciálu proto ustoupily před zaledněním na jih. Od počátku holocénu pak postupně znovu osídlovaly severnější oblasti. Nová území osídlují efektivněji partenogenetické samice, neboť mohou založit novou populaci z pověstného jediného vajíčka, čili jediná samice může dát vznik nové populaci. Partenogenetické populace mohou také vznikat kontinuálně z bisexuálních linií, zatímco opačný proces, vznik bisexuálních populací z partenogenetických, nebyl nikdy doložen a ani se nepředpokládá. Ekologický scénář situaci doplňuje o skutečnost, že bisexuální populace nebyly a dosud nejsou schopny v rámci areálů kompetitivně nahradit populace partenogenetické.
 Rozmanitost reprodukčních způsobů platí ve většině případů hlavně pro sladkovodní lasturnatky evropského kontinentu. U mořských a tropických skupin převažují druhy bisexuální, což je obecně přičítáno vyšší stabilitě tohoto prostředí v holocénu. Jaké přesné mechanizmy k tomuto stavu vedou, je však předmětem úvah spíše hypotetických.
Rozmanitost reprodukčních způsobů platí ve většině případů hlavně pro sladkovodní lasturnatky evropského kontinentu. U mořských a tropických skupin převažují druhy bisexuální, což je obecně přičítáno vyšší stabilitě tohoto prostředí v holocénu. Jaké přesné mechanizmy k tomuto stavu vedou, je však předmětem úvah spíše hypotetických.
Pohlavní a nepohlavní rozmnožování – věčný problém evoluční biologie
Ještě pozoruhodnější než geografická partenogeneze je dlouhodobé přetrvávání nepohlavních linií. V literatuře najdeme teorie, které jejich existenci podporují, stejně jako teorie pesimistické a striktně zamítavé. Narážíme zde totiž na obecný evolučněbiologický problém. Dlouhodobá asexualita (v řádu milionů let) se dostává do rozporu s tvrzením, že druh, který ztratil schopnost pohlavního rozmnožování, je dříve či později nevyhnutelně odsouzen k zániku. Příčinou (teoreticky) nevalných vyhlídek asexuálů je hromadění škodlivých mutací, neschopnost dostatečně pružně reagovat na měnící se prostředí a nižší schopnost čelit neustálému tlaku parazitů a patogenů. Jenže zástupci zmíněné skupiny Darwinulocopina přežívají bez pohlavního rozmnožování celých 245 milionů let! U vířníků skupiny Bdeloidea se délka asexuální existence odhaduje na 40 milionů let. Navíc si prokazatelně vytvořili komplexní fyziologickou obranu proti více než 50 druhům parazitických hub, které je napadají. A zdaleka nejde o poslední přežívající populace zdecimované sexuálními příbuznými. Naopak, např. druh Darwinula stevensoni je rozšířen kosmopolitně a střídmý odhad délky asexuální existence tohoto druhu přitom činí přinejmenším 25 milionů let. Ne všechny striktně partenogenetické lasturnatky jsou kosmopolitní, to je pravda, ale ty, které jsou, mohou žít ve velmi rozmanitých podmínkách. Obecně lze partenogenetické druhy charakterizovat velice rychlou a efektivní reprodukcí, neboť eliminovaly tvorbu samců, kteří se sami nemnoží. Odpovídají také hypotetickému konceptu GPG (general purpose genotype), který se používá zejména při charakterizaci všudypřítomných a těžko likvidovatelných plevelů. Lze tedy říci, že lasturnatky jsou nositeli univerzálního genotypu, resp. genotypu umožňujícího využívat velmi široké spektrum podmínek vnějšího prostředí a zároveň tolerantního k nejrůznějším faktorům.
A zdaleka nejde o poslední přežívající populace zdecimované sexuálními příbuznými. Naopak, např. druh Darwinula stevensoni je rozšířen kosmopolitně a střídmý odhad délky asexuální existence tohoto druhu přitom činí přinejmenším 25 milionů let. Ne všechny striktně partenogenetické lasturnatky jsou kosmopolitní, to je pravda, ale ty, které jsou, mohou žít ve velmi rozmanitých podmínkách. Obecně lze partenogenetické druhy charakterizovat velice rychlou a efektivní reprodukcí, neboť eliminovaly tvorbu samců, kteří se sami nemnoží. Odpovídají také hypotetickému konceptu GPG (general purpose genotype), který se používá zejména při charakterizaci všudypřítomných a těžko likvidovatelných plevelů. Lze tedy říci, že lasturnatky jsou nositeli univerzálního genotypu, resp. genotypu umožňujícího využívat velmi široké spektrum podmínek vnějšího prostředí a zároveň tolerantního k nejrůznějším faktorům.
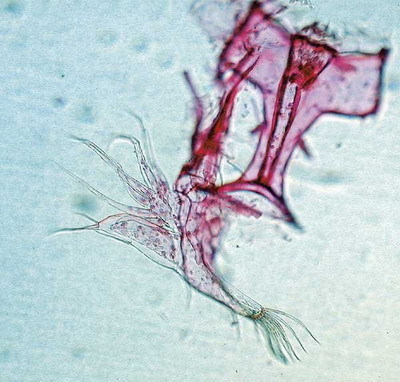 Hypotéza GPG nadále zůstává pouze hypotézou, neboť poněkud koliduje s dalšími teoriemi, které se snaží vysvětit evoluční úspěch dlouhodobých asexuálů. Jednou z nich je např. koncept klonální variability, spatřující úspěch dlouhodobě asexuálních linií v jejich rozrůznění na určitý počet klonů, které vzniká díky hromadění výhodných mutací a meziklonální selekci. Oba mechanizmy podporují vznik specializovaných genotypů adaptovaných na různé typy prostředí.
Hypotéza GPG nadále zůstává pouze hypotézou, neboť poněkud koliduje s dalšími teoriemi, které se snaží vysvětit evoluční úspěch dlouhodobých asexuálů. Jednou z nich je např. koncept klonální variability, spatřující úspěch dlouhodobě asexuálních linií v jejich rozrůznění na určitý počet klonů, které vzniká díky hromadění výhodných mutací a meziklonální selekci. Oba mechanizmy podporují vznik specializovaných genotypů adaptovaných na různé typy prostředí.
Jak je vidět, v problematice nepohlavního rozmnožování zdaleka není jasno (a pravděpodobně ještě dlouho nebude), přestože téma evoluce rozmnožování se v literatuře přemílá již po desetiletí a vznikly desítky hypotéz, proč právě sexuální reprodukce převládá nad asexuální (viz např. článek Jaroslava Flegra, Vesmír 74, 268, 1995/5).
Výhody lasturnatek jako modelu evoluční biologie
A proč jsou lasturnatky natolik unikátním modelem? Důvodů je více. Předně využívají hned několik způsobů reprodukce, které je většinou možné celkem snadno a rychle identifikovat. Mezi lasturnatkami dokonce nacházíme přechod od bisexuální reprodukce k partenogenezi s pravděpodobně nejvyšší frekvencí ze všech živočichů. A proto představují mimořádně významnou modelovou skupinu při studiu selektivních mechanizmů, které udržují u organizmů všeobecně rozšířenou sexuální reprodukci, 4) přestože z ní plynou jasné nevýhody až rizika. Díky své vápenité schránce (karapaxu), jež za vhodných sedimentačních podmínek fosilizuje, poskytují lasturnatky náhled hluboko do minulosti a umožňují zjistit způsob rozmnožování i u populace, která žila před stovkami milionů let. Podobnou kombinaci vlastností najdeme u jiných organizmů těžko. Je pravda, že zjistit pohlaví živočicha, po kterém zbyla kolikrát ani ne milimetrová, často poškozená schránka, není vždy jednoduché. Můžeme se však opřít přinejmenším o tři typy znaků, které při jisté obezřetnosti a opatrnosti fungují bezpečně. První takovou vlastností je velikost karapaxu, přesněji poměr výšky k délce, daný zejména uspořádáním a velikostí vnitřních orgánů. Karapax většiny sladkovodních lasturnatek je v tomto ohledu sexuálně dimorfní. Podle velikosti nebo délko-šířkového indexu bezpečně určíme pohlaví i u silněji poškozeného materiálu. U různých lasturnatek se ovšem vzájemná velikost samců vůči samicím liší. V některých případech vyšší hodnota indexu ukazuje na samčí pohlaví, jindy zas na samičí. U našich skupin většinou platí, že samci jsou subtilnější. Dlouhověcí asexuálové skupiny Darwinulocopina pečují o potomstvo, a tudíž mají na hřbetní straně těla vytvořenou plodovou komůrku, kde se vyvíjejí larvy prvních několika stadií. Její přítomnost se na karapaxu projeví zřetelnou výdutí – další velmi užitečný znak. Karapax je tvořen dvěma miskami – levou a pravou lasturkou – které jsou přibližně uprostřed spojeny svěracími svaly (podobně jako u mlžů). Na vnitřních stranách lasturek se zachovávají otisky po svalových úponech. Jejich pozice je opět dimorfní. U samic dlouhodobě asexuálních linií jsou díky plodové komůrce posunuty zřetelně dopředu. Dalším možným vodítkem k zjištění pohlaví je otisk gonád. Tento znak má však věrohodnou vypovídací hodnotu pouze u velmi dobře zachovaných exemplářů. V každém případě je vždy nezbytné analyzovat pouze karapaxy dospělých jedinců – u mladších vývojových stadií tyto vlastnosti neplatí a vedou k omylům. Naštěstí schránka lasturnatek nese dostatečné množství znaků potřebných k rozlišení mladého jedince od dospělce.
Je pravda, že zjistit pohlaví živočicha, po kterém zbyla kolikrát ani ne milimetrová, často poškozená schránka, není vždy jednoduché. Můžeme se však opřít přinejmenším o tři typy znaků, které při jisté obezřetnosti a opatrnosti fungují bezpečně. První takovou vlastností je velikost karapaxu, přesněji poměr výšky k délce, daný zejména uspořádáním a velikostí vnitřních orgánů. Karapax většiny sladkovodních lasturnatek je v tomto ohledu sexuálně dimorfní. Podle velikosti nebo délko-šířkového indexu bezpečně určíme pohlaví i u silněji poškozeného materiálu. U různých lasturnatek se ovšem vzájemná velikost samců vůči samicím liší. V některých případech vyšší hodnota indexu ukazuje na samčí pohlaví, jindy zas na samičí. U našich skupin většinou platí, že samci jsou subtilnější. Dlouhověcí asexuálové skupiny Darwinulocopina pečují o potomstvo, a tudíž mají na hřbetní straně těla vytvořenou plodovou komůrku, kde se vyvíjejí larvy prvních několika stadií. Její přítomnost se na karapaxu projeví zřetelnou výdutí – další velmi užitečný znak. Karapax je tvořen dvěma miskami – levou a pravou lasturkou – které jsou přibližně uprostřed spojeny svěracími svaly (podobně jako u mlžů). Na vnitřních stranách lasturek se zachovávají otisky po svalových úponech. Jejich pozice je opět dimorfní. U samic dlouhodobě asexuálních linií jsou díky plodové komůrce posunuty zřetelně dopředu. Dalším možným vodítkem k zjištění pohlaví je otisk gonád. Tento znak má však věrohodnou vypovídací hodnotu pouze u velmi dobře zachovaných exemplářů. V každém případě je vždy nezbytné analyzovat pouze karapaxy dospělých jedinců – u mladších vývojových stadií tyto vlastnosti neplatí a vedou k omylům. Naštěstí schránka lasturnatek nese dostatečné množství znaků potřebných k rozlišení mladého jedince od dospělce.
Věřme, že právě studium lasturnatek povede nejen k hlubšímu pochopení příčin a mechanizmů pestré škály reprodukčních způsobů lasturnatek, ale přispěje alespoň nepatrným dílem i k pochopení některých principů evoluční biologie.
Literatura
Chaplin J. A, Havel J. E., Hebert P. D. N.: Sex and ostracods. Tree 9, 435–439, 2003Little T. J., Hebert P. D. N.: Ancient asexuals: scandal or artifact? Tree 11, 296–267, 1996
Martens K. (ed): Sex and Parthenogenesis. Backhuys Publishers, leden 1998
Martens K., Rossetti G., Horne D. J.: How ancient are ancient asexuals? Proc. R. Soc. Lond. B 270, 723–729, 2003
Van Doninck K., Schön I., De Bruyn L., Martens K.: A general purpose genotype in an ancient asexual. Oecologia 132, 205–212, 2002
Poznámky
Ke stažení
 článek ve formátu pdf [465,55 kB]
článek ve formátu pdf [465,55 kB]
O autorovi
Radka Symonová
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
