









Umlčování genů aneb Co dokáže malá RNA
| 14. 4. 2005 Donedávna se myslelo, že DNA a proteiny jsou hlavní hybatelé života a RNA plní většinou jen funkci zprostředkovatele mezi nositelem návodu – DNA – a vykonavatelem – proteinem. A ejhle, v poslední době se ukazuje, že jakési malé RNA mohou ovládat činnost genů umlčováním jejich exprese, a to hned na několika úrovních. Nejnovější poznatky naznačují, že tímto způsobem je kontrolována funkce nejméně třetiny lidských genů.
Donedávna se myslelo, že DNA a proteiny jsou hlavní hybatelé života a RNA plní většinou jen funkci zprostředkovatele mezi nositelem návodu – DNA – a vykonavatelem – proteinem. A ejhle, v poslední době se ukazuje, že jakési malé RNA mohou ovládat činnost genů umlčováním jejich exprese, a to hned na několika úrovních. Nejnovější poznatky naznačují, že tímto způsobem je kontrolována funkce nejméně třetiny lidských genů.
 Sláva kvasinky Saccharomyces cerevisiae jako prvotřídní modelky pro genetická studia pochodů na úrovni eukaryotické buňky je mimo jiné založena na vlastnosti, kterou se kvasinka od vyšších eukaryotických organizmů výrazně liší. Je to přebujelá homologní rekombinace, 1) která dovoluje cílené úpravy včetně úplné eliminace libovolného genu, respektive sekvence DNA. Tím se otvírá brána k systematickému studiu úloh jednotlivých genů a jiných nukleotidových sekvencí v životě kvasinkové buňky. Úplným odstraněním (delecí) genu vytrhneme stromek genového vyjádření (exprese) i s kořeny. Není-li gen, nemůže se pochopitelně projevit ani enzymová, regulační, stavební či jiná aktivita jeho konečného produktu – proteinu nebo RNA. Poškození genu však není jediný způsob jak zabránit jeho vyjádření. Pouť genetické informace od DNA k proteinu je dlouhá, a není tudíž divu, že existuje mnoho míst vhodných k umlčování. Expresi genu lze v eukaryotické buňce přerušit v kterémkoli z jednotlivých kroků procesu (obrázek).
Sláva kvasinky Saccharomyces cerevisiae jako prvotřídní modelky pro genetická studia pochodů na úrovni eukaryotické buňky je mimo jiné založena na vlastnosti, kterou se kvasinka od vyšších eukaryotických organizmů výrazně liší. Je to přebujelá homologní rekombinace, 1) která dovoluje cílené úpravy včetně úplné eliminace libovolného genu, respektive sekvence DNA. Tím se otvírá brána k systematickému studiu úloh jednotlivých genů a jiných nukleotidových sekvencí v životě kvasinkové buňky. Úplným odstraněním (delecí) genu vytrhneme stromek genového vyjádření (exprese) i s kořeny. Není-li gen, nemůže se pochopitelně projevit ani enzymová, regulační, stavební či jiná aktivita jeho konečného produktu – proteinu nebo RNA. Poškození genu však není jediný způsob jak zabránit jeho vyjádření. Pouť genetické informace od DNA k proteinu je dlouhá, a není tudíž divu, že existuje mnoho míst vhodných k umlčování. Expresi genu lze v eukaryotické buňce přerušit v kterémkoli z jednotlivých kroků procesu (obrázek).
 Mezi nejatraktivnější zůsoby jak přerušit tok informace patří RNA-interference (RNAi), která umlčuje vyjádření genu po transkripci, tj. poměrně brzy. Co to znamená? Vpravíme-li určitou dvouřetězcovou RNA (dsRNA) např. do ocásku elegantní hlístice Caenorhabditis elegans (viz Vesmír 82, 133, 2003/3 a 83, 167, 2004/3), umlčíme určitý gen nejen v okolí vpichu, ale také ve vzdálených oblastech, kam interferující RNA spontánně a bez problémů pronikne. Úžasné je to, že na rozdíl od cíleného vytvoření delečního mutanta postupem, který je snadný jen u kvasinek a několika dalších výjimečných tvorů, je umlčování pomocí RNA mnohem univerzálnější. Navíc gen může zůstat nepoškozen, naděje na obnovu jeho exprese tedy přetrvává.
Mezi nejatraktivnější zůsoby jak přerušit tok informace patří RNA-interference (RNAi), která umlčuje vyjádření genu po transkripci, tj. poměrně brzy. Co to znamená? Vpravíme-li určitou dvouřetězcovou RNA (dsRNA) např. do ocásku elegantní hlístice Caenorhabditis elegans (viz Vesmír 82, 133, 2003/3 a 83, 167, 2004/3), umlčíme určitý gen nejen v okolí vpichu, ale také ve vzdálených oblastech, kam interferující RNA spontánně a bez problémů pronikne. Úžasné je to, že na rozdíl od cíleného vytvoření delečního mutanta postupem, který je snadný jen u kvasinek a několika dalších výjimečných tvorů, je umlčování pomocí RNA mnohem univerzálnější. Navíc gen může zůstat nepoškozen, naděje na obnovu jeho exprese tedy přetrvává.
Potranskripční umlčování genů, které se původně považovalo za vzácný jev, se stalo horkým tématem současné molekulární biologie. Schopnost dvouřetězcové nebo vlásenkovité RNA vyvolat destrukci odpovídající homologní mediátorové RNA je dnes jedním z nejdůležitějších nástrojů molekulárních biologů zabývajících se eukaryotickými organizmy. Jde asi o starobylý mechanizmus protivirové obrany, který brání také přeskokům transpozonů, a samozřejmně i o způsob regulace vlastních genů. Umlčování genů při interferenci RNA můžeme charakterizovat:
- účinné agens je dsRNA,
- účinek je vysoce specifický,
- pro rozsáhlý následek stačí pár molekul dsRNA,
- molekuly dsRNA působí alespoň u některých organizmů i na buňky velmi vzdálené od místa vstupu dsRNA do organizmu.
 Objev tohoto jevu pochází z roku 1998. 3) Poprvé byla dsRNA pro umlčení genů destrukcí homologní mRNA použita na Caenorhabditis elegans. Brzy se ukázalo, že tento zázračný mechanizmus lze využít i u dalších eukaryotických organizmů. Dnes se již věří, že dsRNA může umlčovat geny většiny eukaryotických organizmů včetně kvasinek a člověka.
Objev tohoto jevu pochází z roku 1998. 3) Poprvé byla dsRNA pro umlčení genů destrukcí homologní mRNA použita na Caenorhabditis elegans. Brzy se ukázalo, že tento zázračný mechanizmus lze využít i u dalších eukaryotických organizmů. Dnes se již věří, že dsRNA může umlčovat geny většiny eukaryotických organizmů včetně kvasinek a člověka.
 Příprava dsRNA kopie úseku dsDNA in vitro není pro genového inženýra složitým úkolem. V souvislosti s umlčováním se osvědčily např. systémy využívající dvojici bakteriofágových RNA-polymeráz, které rozeznávají promotory např. u genů kolifága T7 nebo T3. Expresní vektor umožňující transkripci in vitro musí obsahovat úsek bohatý na restrikční místa, obklopený promotory T7 a T3 z obou stran (obrázek). Do vybraného restrikčního místa se vloží komplementární DNA (cDNA) odvozená od zvolené mRNA. Takto vzniklá rekombinantní DNA slouží jako matrice pro syntézu obou řetězců dsRNA in vitro. 4) Za vhodných podmínek se oba získané komplementární řetězce spontánně spojí podle zákona komplementarity, a tím vznikne dvouřetězcová RNA. Existují údaje, které ukazují, že určitý nespecifický inhibiční efekt se může dostavit i u genů, které jsou identické jen asi z 80 % v nukleotidové sekvenci.
Příprava dsRNA kopie úseku dsDNA in vitro není pro genového inženýra složitým úkolem. V souvislosti s umlčováním se osvědčily např. systémy využívající dvojici bakteriofágových RNA-polymeráz, které rozeznávají promotory např. u genů kolifága T7 nebo T3. Expresní vektor umožňující transkripci in vitro musí obsahovat úsek bohatý na restrikční místa, obklopený promotory T7 a T3 z obou stran (obrázek). Do vybraného restrikčního místa se vloží komplementární DNA (cDNA) odvozená od zvolené mRNA. Takto vzniklá rekombinantní DNA slouží jako matrice pro syntézu obou řetězců dsRNA in vitro. 4) Za vhodných podmínek se oba získané komplementární řetězce spontánně spojí podle zákona komplementarity, a tím vznikne dvouřetězcová RNA. Existují údaje, které ukazují, že určitý nespecifický inhibiční efekt se může dostavit i u genů, které jsou identické jen asi z 80 % v nukleotidové sekvenci.
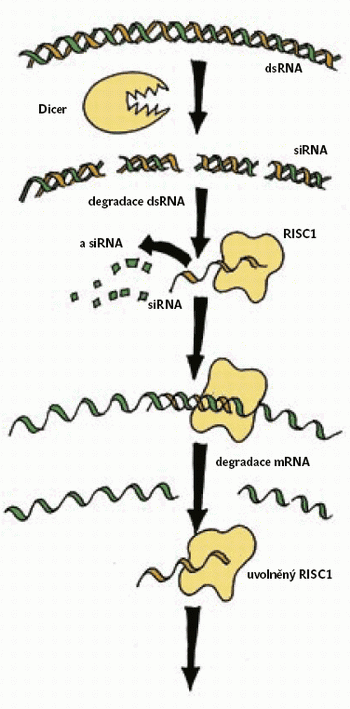
Když vpravíme tuto RNA do organizmu (obrázek), rozštěpí se v buňkách na úseky dlouhé 21–23 nukleotidů (siRNA), a to za pomoci nukleázy zvané Dicer. Potom se vytvářejí tzv. RISC-komplexy 5) těchto krátkých úseků (siRNA) s nukleázami, které nakonec naštěpí mRNA. Při pohledu na schéma (obrázek) snáze pochopíme, proč je tato RNAi považována za způsob potranskripčního umlčování. Množí se však údaje o tom, že dsRNA může působit ještě jinak.
O čem nás poučily kvasinky?
Již dříve jsme si ukázali, že modelová kvasinka Saccharomyces cerevisiae nám díky své zvýšené homologní rekombinaci umožňuje snadno umlčet jakýkoliv gen, a to buď cílenou delecí, nebo cíleným přerušením prostřednictvím vloženého jiného genu. Umlčování genů na základě interference RNA dosahuje obdobného výsledku, tj. ruší funkci genu, a to i v organizmech, které nám neposkytují výhody přebujelé homologní rekombinace. Byly to však opět experimenty s kvasinkami, tentokrát Schizosaccharomyces pombe, které přinesly novýpohled na umlčující účinek dsRNA. Určité oblasti eukaryotických genomů se vyznačují velkým počtem opakování sekvencí nukleotidů. Takové oblasti často přiléhají k centromerám, které hrají rozhodující roli při mitotickém rozchodu chromozomů. Přeskokům transpozonů vložených do těchto oblastí může zabránit vysoce kompaktní chromatin. Vznik tohoto tzv. tichého chromatinu vyžaduje deacetylaci histonu H3 a jeho následnou metylaci na lyzinu. 6)
Určité oblasti eukaryotických genomů se vyznačují velkým počtem opakování sekvencí nukleotidů. Takové oblasti často přiléhají k centromerám, které hrají rozhodující roli při mitotickém rozchodu chromozomů. Přeskokům transpozonů vložených do těchto oblastí může zabránit vysoce kompaktní chromatin. Vznik tohoto tzv. tichého chromatinu vyžaduje deacetylaci histonu H3 a jeho následnou metylaci na lyzinu. 6)
Interference RNA zahrnuje u S. pombe funkce několika složek, které pravděpodobně hrají roli při degradaci dsRNA na siRNA. Jsou to již dříve zmíněná Dicer-nukleáza, protein argonaut a RNA-dependentní RNA–polymeráza (Rpd1). Delece kteréhokoliv z genů kódujících tyto proteiny vede ke ztrátě schopnosti vytvořit tichý chromatin, což svědčí pro to, že interference RNA je součástí širšího mechanizmu, který může dokonce umlčovat geny již na úrovni zahájení transkripce (obrázek). 7)
Umlčování genů jako terapie dědičných chorob
V červenci 2004 se objevila zpráva o úspěšné terapii myši, která strádala spinocerebelární ataxií, dědičnou chorobou, jež je způsobena dominantní mutantní alelou genu SCA1. V důsledku této mutace produkují specifické buňky v mozečku toxický protein způsobující pohybové obtíže. Beverly Davidson a spolupracovníci z Iowské univerzity úspěšně umlčeli mutovaný gen pomocí interference RNA. Interferující RNA (siRNA) působila přímo v mozku pět týdnů staré myši. Po šesti týdnech terapie neměla již myš žádné pozorovatelné obtíže a neobjevily se ani žádné vedlejší účinky.Tyto výsledky naznačují, že by se podobné postupy daly využít i při léčbě některých lidských dědičných chorob. Navíc je pravděpodobné, že techniky umlčování založené na aplikaci dsRNA budou alespoň v některých případech mnohem lepší pro léčení následků genetických poruch než terapie, která je založena na aplikaci DNA.
Poznámky
SLOVNÍČEK
centromera – úsek chromozomu držící pohromadě sesterské chromatidy; úsek DNA pro připojení mikrotubulů mitotického vřeténka (Vesmír 84, 138, 2005/3)delece – ztráta úseku chromozomu
genová exprese, genové vyjádření – celý proces od přečtení genetické informace DNA až po „výrobu“ bílkoviny
promotor – sekvence v DNA, na niž se navazuje RNA-polymeráza, která zajistí transkripci přilehlého genu
rekombinace – proces, při němž jsou chromozomy nebo molekuly DNA rozštěpeny na fragmenty, které jsou znovu pospojovány, ale jinak
dsRNA – dvouřetězcová RNA
mRNA – informační nebo mediátorová RNA řídí vznik bílkoviny v ribozomu, viz heslo translace
RNAi – interference RNA je proces, při kterém dochází k sekvenčně specifickému umlčení genu za pomoci homologní dsRNA
vlásenkovitá RNA – jednořetězcová RNA tvaru vlásenky, má dvě poloviny komplementární, takže vytváří dvojšroubovici spinocerebelární ataxie – dědičné onemocnění míchy a mozečku způsobující těžké poruchy koordinace pohybů, poruchy citlivosti ad.
transkripce – součást genové exprese, která spočívá v kopírování jednoho vlákna DNA do komplementární sekvence RNA za pomoci enzymu RNA-polymerázy, předchází translaci
translace – součást genové exprese, při níž se pořadí nukleotidů v RNA převádí do pořadí aminokyselin v bílkovině, odehrává se na ribozomech, do nichž informaci přináší mediátorová RNA, následuje po transkripci
transpozon – transponovatelný, přemístitelný element; úsek DNA, který se může v rámci genomu přemístit jinam
Ke stažení
 článek ve formátu pdf [605,11 kB]
článek ve formátu pdf [605,11 kB]
O autorovi
Vladimír Vondrejs

Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
