









Morfologická transformace a ontogeneze
| 8. 9. 2003Ať už si o vzniku evolučních novinek myslíme cokoli, jedno víme. Že evoluce morfologických tvarů je vlastně evolucí individuálního vývoje, evolucí ontogeneze, aspoň pokud jde o mnohobuněčné organizmy. Morfologie mnohobuněčného těla vzniká v každé generaci znovu individuálním vývojem; přímo se dědí toliko recept na ontogenezi. Mají vlastně pravdu ti, kdo říkají, že člověk rozhodně nevznikl z opice to jenom z vajíčka, z něhož se dělala opice, vzniklo vajíčko, z něhož se dělá člověk.
Učili jsme se, že ontogeneze opakuje fylogenezi (což říká rekapitulační neboli Haeckelův zákon), a zdá se nám tedy, že k jednoduché ontogenezi předka přidává evoluce další fáze, až vznikne složitější ontogeneze potomka; jinak by samozřejmě dojem rekapitulace vzniknout nemohl. Tak třeba předek byl jednobuněčný, potomek je nejprve jednobuněčný (zygota), posléze mnohobuněčný. Neboli ontogeneze potomka (jednobuněčnost mnohobuněčnost) skutečně v hrubých rysech opakuje jeho evoluci. Jiný konkrétní případ může být ontogeneze platýse. Z jikry se líhne jako celkem normální ryba (i jeho předky byly celkem normální ryby), a teprve později ulehne na bok na mořské dno, zploští se a jeho lebka se tak zkroutí, že spodní oko se jakoby přesune na horní (nikoli ovšem hřbetní!) stranu hlavy. Tato fáze asymetricky zkroucené ryby byla tedy k původní rybí ontogenezi přidána a vzhled dospělého předka platýsů můžeme z ontogeneze dnešního platýse vskutku zhruba odhadnout. Tomuto způsobu evoluce ontogeneze říkáme terminální adice (což znamená, že něco bylo přidáno na konec vývoje).
Terminální adice ovšem zdaleka není jediným způsobem evolučních změn ontogeneze. Primitivní hmyz byl bezkřídlý, s víceméně přímým vývojem (pouze rozstříhaným na několik period pravidelným svlékáním kutikuly, přičemž jednotlivé úseky, instary, se od sebe téměř nelišily). Okřídlený dospělec (u různých vymřelých skupin bylo několik okřídlených instarů, v dnešní fauně mají jepice dva a veškeré ostatní skupiny jediný okřídlený instar) byl k této ontogenezi skutečně jakoby přidán. Když ale později přibylo další stadium před dospělce, totiž kukla, nešlo jistě o terminální adici. U hmyzu skupiny Holometabola (brouků, motýlů, much a podobně) totiž proběhla výrazná ekologická a morfologická specializace larev k jiným způsobům života, než vede dospělec (srovnejte housenku a motýla nebo muší larvu s dospělou mouchou), přičemž součástí této specializace byl i zánik vnějších základů křídel u larev. Jestliže primitivní hmyz má nelétavé larvy s postupně narůstajícími nepohyblivými výběžky, z nichž se jednou u dospělce stanou funkční křídla, larvy holometabolního hmyzu žádné vnější základy křídel nemají a křídlo vzniká mnohem prudší přeměnou. Kukla nežere a obyčejně se ani nehýbá a cele se věnuje pouze náročné metamorfóze; na vznik křídel je tu jediný specializovaný instar. 1) Toto stadium bylo do ontogeneze jaksi přidáno, ale doprostřed ontogenetické dráhy. Jenže ani housenka, tedy specializovaná larva hmyzu, už schématu rekapitulace neodpovídá, neboť žádný předek motýlů v dospělosti nevypadal jako housenka.
Heterochronie
Abychom mohli hovořit o ontogenezi jako o sledu oddělených stadií, musíme používat hodně hrubé zjednodušení. Ve skutečnosti je samozřejmě ontogeneze každého druhu kontinuální proces. Dokonce i ontogeneze hmyzu, byť jakoby rozdělená do izolovaných instarů, je zcela povlovná: to, co se svléká, zahazuje, a tedy skokově mění, je kutikula, ale kutikula není zvíře, kutikula je sekret produkovaný pokožkovými buňkami, tedy věc de facto neživá. Přesto ještě chvíli setrvejme u formální představy ontogeneze jako sekvence stadií: evoluce pak může spočívat i ve změnách časování jednotlivých klíčových okamžiků této sekvence. Takzvaná heterochronická evoluční změna spočívá v tom, že se změní relativní rychlost, načasování začátku nebo načasování konce růstu nějaké tkáně či orgánu. Zatímco samotná změna ontogeneze nemusí být příliš nápadná, morfologický (a ekologický) výsledek může být dosti zásadní.
Podívejme se na vztah pohlavních buněk (neboli gamet) a buněk somatických, tedy všech buněk tkání a orgánů, které se nebudou přímo podílet na vzniku nové generace. U mnoha organizmů tvrdé rozdělení pohlavních a somatických buněk neexistuje a nová generace může vznikat i ze somatické buňky (vegetativní množení rostlin i mnoha živočichů), ale stejně je významné zjistit, v kterém okamžiku vývoje somatických tkání začne být organizmus schopen rozmnožování, tedy v kterém okamžiku má funkční zárodečné buňky. Po tomto okamžiku totiž další morfogeneze somatických tkání obvykle končí; značná část organizmů se množí jenom jednou za život, a poté hynou a i ti ostatní po dosažení věku prvního potenciálního rozmnožení spíš stárnou a chátrají, než že by se ještě nějak významně měnili. Což vidíme na sobě.
Heterochronii tedy můžeme chápat jako změnu okamžiku, kdy se organizmus množí, ve srovnání s průběhem ontogeneze jeho těla. Zvlášť významné jsou změny, kdy se odvozený druh začne množit (relativně) dřív, než se množil jeho předek. V tom případě se ontogeneze potomka jaksi zasekne, skončí na ranější fázi somatického vývoje, nedoběhne. Jako by se množilo mládě či larva, tedy nedospělý jedinec. Všimněte si, že zde neplatí rekapitulační zákon; nové dospělé stadium nebylo přidáno k původní ontogenezi, nýbrž naopak, původní koncové stadium bylo ubráno, ontogeneze (formálně vzato) zjednodušena.
Známe dva základní typy heterochronické evoluce. Může dojít ke ztrátě koncového stadia ontogeneze buď relativním zrychlením vývoje zárodečných buněk, anebo relativním zpomalením vývoje ostatních tkání. Morfologický výsledek, tedy zachování juvenilních či larválních znaků v dospělosti, se nazývá pedomorfóza. Asi nejslavnějším příkladem pedomorfózy je axolotl, který se běžně množí jako vodní larva, ale který je za určitých podmínek schopen dokončit kompletní vývoj (tedy změnit se v suchozemského mloka); pedomorfóza mnohých druhů mloků je ovšem pevně fixována a návrat k původnímu složitějšímu vývoji už není možný. Za všechny příklady zmiňme známého macaráta jeskynního (Proteus anguinus). Zároveň existují také dva opačné procesy, totiž relativní zrychlení vývoje tělních buněk a relativní zpomalení vývoje buněk zárodečných, směřující k výsledku zvanému peramorfóza, tedy k takovému odvozenému typu vývoje, který rekapituluje ontogenezi předka, a navíc k ní cosi dodává, a to pozdní stadium nepodobné dospělému stadiu předka. Mezi peramorfní změny patří především různé případy gigantizmu, třeba extrémní zvětšení paroží u některých jelenů.
Člověk je ve srovnání s ostatními primáty jakési přerostlé mládě, či spíše pozdní embryo; dospělý člověk je nápadně podobnější novorozeněti než dospělý šimpanz šimpanzímu novorozenci. Už ve dvacátých letech 20. století byl předložen první seznam údajně pedomorfních znaků člověka: novorození lidé a šimpanzi i dospělí lidé se navzájem podobají v takových vlastnostech, jako je redukované ochlupení, velký mozek, plochý obličej s krátkými čelistmi, umístění velkého lebečního otvoru (a tedy tvar napojení lebky a páteře), stavba rukou a chodidel, jakož i zvědavost, kreativita a smysl pro humor. Do vědecké diskuse vstoupila představa člověka jako pedomorfního lidoopa především zásluhou či vinou Stephena Goulda. Ten ale svou koncepci nepostavil na seznamu znaků, které lze interpretovat jako heterochronické, nýbrž na představě celkového zpomalení vývoje somatických tkání a orgánů. Člověk by tudíž měl být všeobecně pedomorfní primát, ačkoli jednotlivé znaky, například prodloužení zadních končetin, pozdě ukončený růst těla a zpoždění sexuální zralosti, poukazují spíše na změny peramorfní.
Hlavně jsou ale mnohé rysy ontogeneze člověka od ontogeneze ostatních lidoopů odlišné jinak, než aby tento rozdíl bylo možno vysvětlit heterochronickou změnou. Tak například lidský charakter lebeční báze, údajně pedomorfní, je spojen s přestavbou horních cest dýchacích a s poklesem hrtanu na úroveň nižší než u jiných savců (což souvisí se vznikem řeči). Tato změna tedy není nijak evidentně heterochronická. Také lidský mozek není pouze přerostlý mozek šimpanzí, ačkoli mnohé vlastnosti našeho mozku, třeba stupeň rýhování nebo velikost neokortexu, takovou interpretaci umožňují. Zvýrazněnou pravolevou asymetrii lidského mozku nebo vznik Brocova centra v levé hemisféře však lze jen těžko vysvětlit globální pedomorfózou člověka. 2)
Heterochronie a evoluce
Jaký je skutečný význam heterochronických změn v evoluci, to nevíme. Je třeba zdůraznit, že koncepce heterochronií je založena na formálním, geometrickém, v podstatě nebiologickém pohledu na ontogenezi a její eventuální změny. To, že nová morfologická či ekologická forma vznikla jako důsledek heterochronického posunu, neříká nic o tom, zda se na této evoluci podílela selekce či nikoli. Ač je v literatuře patrný jistý sklon interpretovat evoluci heterochronií jako protiklad evoluce přirozeným výběrem, jde o zjevné nedorozumění. Heterochronie je jedno z možných vysvětlení, jak se dělají evoluční novinky, zatímco selekce vysvětluje, proč byly vlastně potřeba.
Tak například axolotl řeší pedomorfózou závažný problém života v horských či pouštních jezerech. Také mláděcí vzhled člověka (věřme, že jde mimo jiné i o kombinaci komplikovaných heterochronií) může být odpovědí na selekční objednávku. Pokud výchova lidského mláděte trvá velmi dlouho a vyžaduje extrémně náročné rodičovské angažmá, je selekčně výhodné vybírat si partnerku co nejmladší, čímž zvyšujeme šanci, že mládě dochová (výběr co nejmladšího muže je zaprvé méně významný vzhledem k menší otcovské investici do potomstva a zadruhé je ve výrazném konfliktu s výběrem partnera co nejbohatšího a nejmocnějšího, který těmito vlastnostmi urovná potomkovi cestu životem, i když třeba sám záhy umře). V dnešní civilizované společnosti každý ví, kdy se narodil, a i my ostatní se to o něm snadno dozvíme nahlédnutím do úředních lejster. Ve společnosti, kde dnešní vývojový cyklus člověka vznikal, neměl nikdo s matrikami bližší zkušenost a mládí nezbývalo než odhadovat z toho, jak potenciální partnerka vypadá. Dodnes jsou vlastnosti, které u žen oceňujeme jako krásné (oči, rty, pleť, postava), v podstatě pevně svázány s mládím a bez invazivního vměšování moderního průmyslu obtížně přežívají třicítku. Výběr partnerek, které vypadají mladě, jistě vedl k postupnému omlazování vzhledu. Dvě ženy hledající ve věku pětadvaceti let partnera (a geny těchto žen) budou mít různý úspěch, vypadá-li jedna na dvacet a druhá na třicet. Že touto selekcí mladých partnerek došlo k omlazení obou pohlaví, není žádná záhada máme společné a stále se míchající geny.
Jiným příkladem adaptivní heterochronické změny mohou být pomocníci (helpers) sociálně žijících živočichů, pohlavně dospělí jedinci, kteří by se mohli samostatně rozmnožovat, a přece raději pomáhají svým rodičům s výchovou svých sourozenců. Jak dlouho bude mladý jedinec pomáhat rodičům, záleží mimo jiné i na riziku případného osamostatnění. V čím větších koloniích jedinců příslušný druh žije, tím menší je naděje, že se vzpurnému jedinci podaří novou úspěšnou kolonii založit, a tím silnější je tlak, aby zůstal doma a pomáhal otci a matce, zvláště když na tom geneticky nic netratí. Představíme-li si situaci, kdy většina pomocníků umře dřív, než se stačí osamostatnit, vyjde nám kolonie nemnožících se jedinců pomáhajících jedinému množícímu se páru či harému tedy právě to, čemu říkáme eusociální uspořádání u termitů nebo několika druhů afrických hlodavců rypošů (Bathyergidae). 3) Rozdíl mezi krátkodobou rodinnou výpomocí floridských sojek křovinných (Aphelocoma coerulescens), které se nijak zvlášť nedivíme, a mezi eusocialitou termitů, kterou složitě vysvětlujeme, pak spočívá pouze v načasování osamostatňovacího kroku.
S koncepcí heterochronií je jeden problém. Obvykle nemáme žádné proximativní 4) biologické vysvětlení, jak k takovým změnám dochází. Známe několik případů heterochronických genů, které mění časování různých událostí v larválním vývoji hlístic; 5) víme, že tyto geny jsou rozšířeny i u jiných živočichů, a leccos nasvědčuje tomu, že i tam tyto geny mění časovou regulaci vývoje. Nejde ovšem o velké evoluční změny. I hlístice s mutantním heterochronickým genem vypadá úplně jako hlístice. Protože nás primárně zajímá podíl heterochronie na vzniku nových tělních plánů a životních strategií, na vzniku nových velkých kladů, musíme jako obvykle v evoluční biologii extrapolovat z malých a dobře dokumentovaných změn k změnám velkým a věřit, že taková extrapolace má smysl. Případ axolotlů, kdy máme formy s kompletní metamorfózou i formy pedomorfní a experimentálně dokážeme měnit jednu v druhou, je zcela výjimečný. Právě v tomto případě vidíme, jak nepatrné události vyvolávají podstatné změny evoluční. Aniž by muselo docházet k velkým a pravděpodobně zhoubným mutacím, může skupina organizmů cestou heterochronické evoluce jaksi uniknout z dosavadní specializace a začít se vyvíjet jiným směrem.
Pokud k tomu ovšem vůbec dochází. Seznam skupin, o nichž někdy někdo prohlásil, že vznikly heterochronickou změnou (obvykle pedomorfně), je totiž dlouhý, ale většinou jde o zbytky předfylogenetických úvah, nebo byly tyto hypotézy vyvráceny zdokonalenými znalostmi o příbuzenských vztazích mezi organizmy. Například mnohonožky se líhnou z vajíčka jako šestinohé larvičky a postupně jim přirůstají další články a končetiny; novorozená mnohonožka tedy vypadá jako hmyz, a hmyz by mohl vzniknout z pedomorfních mnohonožek. Ale nevznikl; mnohonožky jsou hmyzu vzdálenější než korýši.6)
Jsou obratlovci pedomorfní?
Asi nejvýznamnějším aspoň pro nás lidi příkladem možného vzniku velkého kladu cestou pedomorfózy jsou obratlovci. Teorie, že obratlovci (a kopinatci) vznikli z pedomorfních pláštěnců, vychází z nápadné podobnosti obratlovců, kopinatců a larev pláštěnců: jde o volně pohyblivá zvířata s hřbetní strunou a hřbetní nervovou trubicí, zatímco typický dospělý pláštěnec, sumka, je válcovitý přisedlý útvar s nápadně zjednodušenou anatomií, překrytý celulózním pláštěm a věnující se toliko filtrování mořské vody.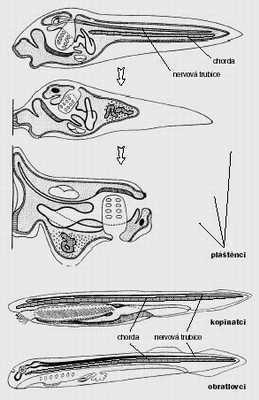
Obratlovci a kopinatci tedy evidentně nemají nic společného s dospělou sumkou, zato s její larvou ano. Ale pozor. Podle stejné logiky totiž můžeme prohlásit, že normální ryba, třeba pstruh, je vlastně pedomorfní platýs. Také mládě platýse je normální ryba podobná pstruhovi, zatímco dospělý platýs leží nesmyslně na boku s oběma očima na jedné tváři. Kupodivu, tato divoká myšlenka o evoluci pstruhů nikdy nikoho nenapadla. Všem je totiž jasné, že mládě platýse není larva a že to, co je na platýsovi opravdu divné a pozoruhodné, není mládě, nýbrž právě ten zvrhlý dospělec.
Abychom mohli předpokládat, že obratlovci jsou pedomorfní pláštěnci, museli bychom uvěřit, že volně pohyblivé stadium pláštěnců je opravdu larva (jak se jí také v učebnicích běžně říká), že přisedlá dospělá sumka odpovídá dospělému stadiu primitivních strunatců a že nejde (jako u dospělého platýse) o apomorfní dodatek k původně jednoduché ontogenezi. Uvěřit tomu všemu je dosti obtížné, přesto bývá každý strunatec, když se hýbá a vypadá jako zvíře, neoblomně označen za pedomorfního. Mnohem jednodušší (a tedy i oprávněnější) intepretace je ovšem taková, že kopinatci a obratlovci jsou podobní larvám sumek proto, že takhle vypadali primitivní strunatci a že tu skutečně odvozenou formu strunatců představuje dospělá sumka. Dospělá sumka vznikla terminální adicí a není divu, že ve své ontogenezi rekapituluje předchozí fylogenezi: rané stadium vývoje sumky vypadá jako dospělý primitivní strunatec.
Představa, že obratlovci jsou pedomorfní, tak vlastně vychází z absurdního nedorozumění o evoluci představující jakýsi lineární žebřík směřující k vrcholu tvorstva, totiž k nám v takovém případě by vskutku každý znak, kterým se obratlovec liší od libovolného neobratlovce, musel být apomorfií obratlovce. Ve skutečnosti jsou samozřejmě obratlovci stejně jako každá jiná skupina mozaikou primitivních a odvozených vlastností. Člověk je v mnohém primitivnější než sumka, životní cyklus člověka je primitivně jednodušší než cyklus sumky, a žádné heterochronické vysvětlení tu k ničemu nepotřebujeme (leda jako vysvětlení toho, proč se sumka nerozmnožuje ve stadiu pohyblivého zvířete, ale až později). 7)
V současnosti upřímně řečeno neexistuje jediná velká skupina organizmů na úrovni třídy nebo kmene, o níž bychom mohli věrohodně prohlásit, že vznikla heterochronicky. Jisté naděje vzbuzují snad jenom ploštěnci a vířníci. Známe několik dobře dokumentovaných případů heterochronie na úrovní druhů či populací, spoustu dalších evolučních událostí umíme s podezřelou lehkostí označit za pravděpodobně heterochronické, dokážeme si představit, jak snadná by byla evoluce nových kmenů pomocí heterochronie ale neznáme jediný věrohodný případ, kdy by pomocí heterochronie nějaký kmen opravdu spolehlivě vznikl.
Poznámky
Ke stažení
 Článek ve formátu PDF [265,07 kB]
Článek ve formátu PDF [265,07 kB]
O autorech
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
