










Evoluce bez adaptací
Evoluce jako soupeření fylogenetických linií
Každá učebnice biologie, když dorazí k evoluci, obsahuje obrázek evoluční řady koní, slonů či lidí. Jak známo, koně se během evoluce zvětšovali, ubývalo jim prstů a měnily se jim stoličky, jak se v důsledku změny prostředí přesouvali z vlhkých pralesů do savan a stepí.
Co ale taková evoluční řada vlastně znamená? Všichni ji patrně budeme vnímat prizmatem přirozeného výběru. V populaci malých a pětiprstých prakoníků se objevil větší či máloprstý mutant, který by v původním lesním prostředí neměl šanci, ale jak Severní Amerika vysychala, mutant se ukázal v konkurenci o zdroje úspěšnějším a primitivní typ postupně nahradil. Zvětšení koní si tedy představujeme jako čistě anagenetický 1) trend, jako důsledek vnitrodruhové soutěže individuálních koní různých typů, jako výsledek normálního přirozeného výběru. Ten by ovšem musel pracovat v nápadně širokém časovém měřítku více než 50 milionů let, aniž by jeho činnost byla přerušena kladogenetickou 2) událostí, speciací. Jakmile totiž dojde k speciaci, selekce začne pracovat izolovaně uvnitř každého z nových druhů, tedy v novém populačním a obvykle i ekologickém kontextu.
Kardinální otázka zní, zda můžeme takový dlouhodobý trend opravdu empiricky potvrdit výzkumem fosilního záznamu a nevznikl-li pouze extrapolací jednotlivých nálezů. Z existence starého malého koně a mladého velkého koně můžeme vyvodit zvětšovací trend, ale klidně se můžeme mýlit, pokud starší nález nepředstavuje předka mladšího nálezu ani něco tomuto předkovi blízkého. To, co bychom tedy potřebovali, je dlouhodobý kontinuální fosilní záznam, kde bychom případně viděli postupné (graduální) změny. Takový záznam je ovšem tak vzácný (a zcela výjimečně se týká nějaké skupiny, jejíž evoluce nás opravdu zajímá), že zjevnou absenci graduálních změn těžko interpretovat.
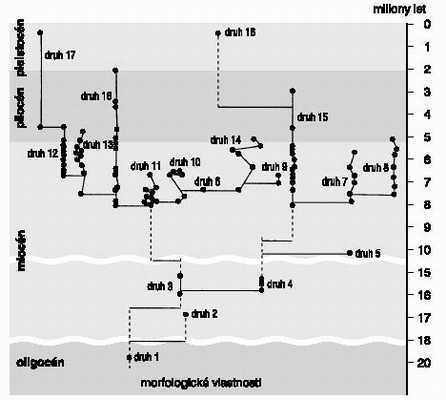
Ještě před několika desetiletími byla interpretace neúplného fosilního záznamu jednoznačně gradualistická. Nalezené fosilie se prostě spojily čárou, a bylo to: starší fosilie byla interpretována jako předek fosilie mladší. Z té doby také víme, že v evoluci obratlovců existují různé fylogenetické zákony (například skoro všichni savci se zvětšují); ono je také těžko neobjevit nějaký ten obecný zákon, hledáme-li jej metodou prokládání přímky dvěma body. Skupina paleontologů kolem Stephena Goulda a Nilese Eldredge předložila počátkem sedmdesátých let 20. století alternativní pohled na skutečnost, že graduální změny nejsou skoro nikde vidět. Tento sám o sobě nepochybný fakt býval vysvětlován nekompletností fosilního záznamu: i kdyby byla evoluce opravdu dokonale povlovná, stejně by z ní zbyly jenom izolované body. Jenže to, co vidíme tam, kde fosilní záznam umožňuje aspoň nějaké závěry, bývá dlouhodobá existence druhu, s nímž se nic moc neděje, načež tento druh zmizí a je nahrazen druhem blízce příbuzným či několika takovými druhy. Gould a Eldredge tvrdí, že druhy během své nudné existence nejsou sice úplně neměnné, ale nejsou na nich patrné žádné trendy jejich morfologické hodnoty nějak chaoticky oscilují kolem počátečního stavu. Významné morfologické (a jistě i ekologické) změny jako by byly spojeny s kladogenezí, s rozštěpením starého druhu do druhů nových. Jak takový fosilní záznam obvykle vypadá, ukazuje obrázek evolučních vztahů osmnácti druhů mořských mechovek rodu Metrarabdotus za posledních 20 000 000 let: nic nenasvědčuje tomu, že by se tu v rámci druhů něco dělo, i když mnohé druhy se ve fosilním záznamu dokázaly udržet po několik milionů let. Vznik morfologických rozdílů mezi jednotlivými druhy je zřejmě spojen s kladogenetickými událostmi.
Gouldově a Eldredgově teorii se dostalo názvu teorie přerušovaných rovnováh (punctuated equilibria) a obecného nepochopení. Především nejde o teorii kreacionistickou. To, co se odehrává v okamžicích zlomu, není žádný tvůrčí akt, nýbrž pouhá alopatrická speciace. 3) Nedostatek přechodných článků je způsoben tím, že k evoluci novinek dochází v malých a obtížně objevitelných populacích někde na periferii areálu původního druhu, tedy někde jinde než v oblasti, kde jsme něco vykopali, a nový druh se na původní území může dostat leda dodatečnou imigrací a objeví se tedy náhle. I zastánci přerušovaných rovnováh považují neúplnost fosilií za důležitý aspekt, ale spíš fragmentárnost prostorovou než časovou. Zadruhé teorie přerušovaných rovnováh nezbytně nepředpokládá velké a náhlé skokové změny: změny probíhající v okamžiku zlomu nemusejí být větší než mutace, které předpokládají gradualisté. Rozdíl je v časovém rozložení evolučních událostí, ne v jejich velikosti. Kde gradualisté předpokládají víceméně rovnoměrný příkon mutací na jednotku času, tam punktuacionisté vidí okamžiky evoluční změny prokládané dlouhými období evoluční nudy, stazigeneze. Kde se stazigeneze bere, není jasné. Může jít o výsledek stabilizující selekce (organizmy se nemění proto, že jsou lépe adaptovány na své prostředí než noví mutanti) i o omezení vznikající variability (organizmy se nemění proto, že se z genetických či embryologických důvodů měnit nemohou a selekce tu nemá s čím pracovat).
Především ale teorie přerušovaných rovnováh ukazuje, že anageneze a kladogeneze nejsou nutně dva nezávislé jevy, že se anagenetické změny mohou koncentrovat do okamžiků speciačních událostí. Anageneze je pak vlastně jakousi vedlejší zplodinou kladogeneze. Když se mateřský druh rozpadne, v každém dceřiném druhu se zafixují jiné evoluční změny, ať už adaptivní nebo náhodné, dosavadní adaptivní historie se jaksi zresetuje, mutace, které už se třeba v mateřském druhu skoro zafixovaly dlouhodobou předchozí selekcí, z nového dceřiného druhu náhle zmizí, protože se třeba odštěpili zrovna ti jedinci, kteří je neměli a začíná se do jisté míry znova.
Konflikt mezi gradualisty a zastánci přerušovaných rovnováh trvá. Empirický paleontologický výzkum nám toho moc neřekne, protože fosilní záznam je všeobecně mizerný. Najdeme-li jasné přerušení rovnováhy, označí to gradualisté za důsledek imigrace jiných druhů odjinud, kde už dávno existovaly, a konkurenčního vytlačení původního druhu. Najdeme-li graduální změnu, řeknou nám punktuacionisté, že jsme zrovna objevili kousek stazigenetického úseku, a protože jej zkoumáme příliš jemnými nástroji, připadá nám to jako trend. Protože nadto jen málokdo pochybuje o tom, že reálně se ve fylogenezi organizmů projevily jak graduální, tak punktuační změny, je jasné, že spor se vede o to, jak jsou časté, a to nám paleontologie opravdu stěží řekne. Jediné, co je těžko zpochybnitelné (a opravdu nápadné), je existence dlouhých evolučních stází.
Zajímavější je tedy podívat se na celou problematiku z biologického hlediska. Představa, že evoluční změny jsou pevně spojeny s okamžiky kladogenetického štěpení, je v dobrém souladu s představami populační genetiky. Jen opravdu dobré alely soutěžící s opravdu špatnými konkurentkami mohou doufat, že se prosadí ve velké populaci poctivým přirozeným výběrem. Zafixovat se náhodně, genetickým driftem, je ve velké populaci úkol na hodně dlouhou dobu a s nejistým výsledkem. Naopak v malé populaci je vysoká šance prosadit se náhodně a selekce v malé populaci pracuje s příbuznými jedinci s obdobnými alelami v obdobném genomovém kontextu. Jenže malé populace jsou fenoménem víceméně efemérním. Malá populace dřív nebo později buď zanikne, anebo vplyne zpět do sousední velké populace a její lokální znamenitosti se opět rozplynou v širokém genofondu druhu. Jestliže ovšem nedojde ke speciaci, která zmrazí malou populaci ve stavu, jehož dosáhla, a umožní jí vyvíjet se jiným směrem než sousední populace, které se náhle právě proto, že proběhla speciace staly geneticky irelevantními.
Dejme tomu. Ale kde se v tom případě berou ty dlouhodobé trendy? Pokud se koně opravdu zvětšují (a oni se zvětšují), není to proto, že malí jedinci víc umírají a hůř se množí, neboť uvnitř druhů se vlastně nic neděje; je to možná proto, že druhy s malými jedinci víc vymírají nebo hůř speciují, popř. obojí. Rozpadne-li se druh, jehož jedinci jsou střední velikosti, na dva dceřiné druhy, jeden menší a druhý větší (uvozovky proto, že nemluvíme o velikosti druhů, tedy o počtu jejich jedinců, ale o velikosti těla těchto jedinců), průměrná velikost zůstává zachována (na obrázku na s. 646 vlevo). Pokud se ale větší druhy budou štěpit častěji, průměrná velikost kladu se bude zvětšovat (na tomtéž obr. obrázek). Nalezneme tudíž trend zvětšování velikosti těla, aniž by se velcí a malí jedinci vůbec museli potkat jako konkurenti v rámci jednoho druhu a aniž by ti velcí museli nějak projevit svou individuální vícecennost. V takovém případě by byl zdánlivě anagenetický trend ve velikosti těla koní určen asymetrickou kladogenezí, procesem, kterému říkáme druhový výběr (species selection).

Něco podobného se ve fosilním záznamu skutečně podařilo prokázat, nikoli ovšem u koní, ale u mořských mlžů. U nich se neprojevují žádné trendy ve změnách velikosti, ba právě naopak, během evoluce zůstává rozložení jejich velikostí po miliony let překvapivě stabilní, a dokonce nezávislé na zeměpisné šířce: všude a vždycky má většina druhů stejně středně velké schránky a druhy hodně velké, nebo naopak malé jsou mnohem vzácnější. Vypadá to, že ta nejčastější velikostní kategorie je zároveň nejvýhodnější, poněvadž menší jedinci mají relativně velké energetické nároky na jednotku hmotnosti a velkým jedincům zas dlouho trvá, než vyrostou. Jenže kupodivu anagenetické změny v rámci jednotlivých linií vůbec nemají tendenci k onomu optimu směřovat, ba dokonce ani v něm zůstávat: velké druhy se klidně dál postupně zvětšují, malé zmenšují a ony středně velké se vyvíjejí vcelku libovolným směrem. Za udržování stabilního rozložení tělesné velikosti mezi druhy je odpovědná pouze rozdílná rychlost vymírání a speciace jednotlivých linií; přirozený výběr mezi jedinci k žádnému optimu nesměřuje.

Některé klady mají tedy tendenci vytvářet víc druhů než jiné. Na evoluci se můžeme dívat nejen jako na konkurenci mezi jedinci v rámci druhu o to, kdo vyprodukuje víc potomstva, ale i na úrovni druhového výběru jako na konkurenci mezi druhy či klady, který z nich vyprodukuje nejvíc dceřiných druhů/kladů. Umožňuje nám pochopit, proč jsou některé skupiny, třeba hmyz, bohatší než jiné. Ukázali jsme si, že kladogeneze je v principu odlišný proces od anageneze, a že tedy k vysvětlování diverzity nám nepostačí princip přirozeného výběru, jakkoli se vznikem druhů určitě také souvisí. Proto se musíme ptát nikoli na to, které znaky umožňují přežití a rozmnožení jedinců, ale spíše na to, které ovlivňují pravděpodobnost speciace nebo vymření v daném kladu.
Popravdě řečeno, většinou jde o stejné znaky. Klady s největší diverzitou jsou ty, které mají rychlý populační růst (daný třeba krátkou generační dobou) a vysokou schopnost využívat různé zdroje (danou buď malou mírou specializace, nebo vysokou mobilitou). To jsou patrně vlastnosti, které jsou prospěšné jak jedincům, tak druhům, poněvadž u nich snižují pravděpodobnost vymření. Druhy s velkou rychlostí rozmnožování mohou dosáhnout (a dosahují) větších populačních početností, mobilita a nízká specializovanost zaručují obsazení většího areálu a obě vlastnosti podporují navíc schopnost dostat se z krizí (rychlým namnožením či emigrací). V těchto případech můžeme mluvit o úspěšnosti daného kladu, aniž ji musíme nutně odlišovat od úspěšnosti jedinců; třeba schopnost aktivního letu je výhodná jak ve vnitrodruhové konkurenci, tak ve vytváření nových kladů, protože snižuje pravděpodobnost vymření. Také proto jsou patrně létající klady tak relativně početné hmyz globálně, ptáci v rámci suchozemských obratlovců, netopýři v rámci savců.
Občas se tvrdí, že nikoli široká tolerance k různým typům prostředí, ale naopak úzká ekologická specializace způsobuje větší radiaci. Africké antilopy ze skupiny Alcelaphini, která zahrnuje buvolce a pakoně, byly vždy druhově rozrůzněnější než impaly (Aepycerotini). To se přisuzuje jejich mnohem větší specializovanosti. Buvolci a pakoně mají specifické nároky na potravu a prostředí, a tak se jejich populace vyskytují ostrůvkovitě, podle nabídky potravních zdrojů. Rozdrobení jejich areálů by mohlo být důvodem častější alopatrické speciace, zatímco u impaly, která je nespecializovaná a žije úplně všude, ke speciaci hned tak nedojde. U impaly ovšem na druhou stranu byla vždy menší pravděpodobnost vymření, takže celková čistá rychlost vytváření nových druhů (speciace minus vymírání) zůstávala v obou skupinách zhruba srovnatelná, i když celkové druhové bohatství nikoli. Ani další příklad, který se uvádí jako doklad druhového výběru prostřednictvím rozdílné specializovanosti, není úplně bez chyb. Mořští plži s planktonními larvami jsou druhově chudší než plži, kteří jsou po celý život přisedlí a vázaní na určité prostředí. Planktonní larvy brání izolaci jednotlivých populací, a tak snižují pravděpodobnost alopatrické speciace. Jenže když se podrobně podíváme na fylogenezi nějaké větší skupiny mořských plžů, zjistíme, že přechod od planktonního k přisedlému způsobu larválního života je nesrovnatelně pravděpodobnější než opačný postup (neboť opačně se to zřejmě nestalo nikdy). Není pak divu, že přisedlí jsou druhově bohatší, když přisedlosti během fylogeneze jenom přibývá, bez ohledu na to, jaký je reálný vztah rychlosti speciace k přítomnosti pohyblivých larev. Zatím tedy neexistuje dobrý doklad, že by specializace zvyšovala diverzitu prostřednictvím větší pravděpodobnosti geografické izolace a následné alopatrické speciace.
Všechny znaky, o nichž jsme zatím mluvili, jsou kromě toho, že mohou podnítit častější odštěpování nových linií, také nějak výhodné z hlediska jedince, a jejich výskyt tedy lze vysvětlovat i klasicky darwinovsky. Druhový výběr ovšem začne být zajímavý tehdy, když znaky, které vedou k častějšímu odštěpování nových linií, nejsou zároveň nijak adaptivní v darwinovském slova smyslu, či jsou dokonce darwinovsky škodlivé. Moc jich neznáme, poněvadž se špatně hledají. Nemohou totiž souviset s pravděpodobností vymření (to by byly asi zároveň individuálně adaptivní), ale s rychlostí speciace, a my moc neznáme faktory, které jsou ve speciaci skutečně rozhodující. Pár vhodných kandidátů by se ale našlo. Třeba ten už zmíněný barevný dimorfizmus a ornamentace ptáků, které korelují s druhovým bohatstvím zřejmě díky vztahu k pohlavnímu výběru. Barevní ptáci těžko přežívají líp než ptáci nenápadně zbarvení, takže tento znak asi opravdu jde proti individuální selekci. U jiných organizmů by člověk předpokládal, že nápadné zbarvení může být výstražné, tedy signalizující predátorovi Nežer mě, jsem nejedlý nebo nebezpečný! (jako v případě vos, mloků nebo skunků), ale ptáci jsou až na zanedbatelné výjimky natolik poživatelní, že by to pro ně nebyl dobrý způsob ochrany. Na druhou stranu barevnost ptáků není výhodná pouze z hlediska výběru druhového (neboť vede k vyšší rychlosti speciace), ale i z hlediska jedince, jelikož pohlavní výběr rozhoduje o reprodukční zdatnosti samců. Čili k vysvětlení barevné diverzity ptáků možná vůbec nepotřebujeme druhový výběr a vystačíme si s výběrem pohlavním.
V každém případě existuje vzrušující možnost, byť zatím neprokázaná, že některé znaky tu nejsou proto, že umožnily lepší přežití a rozmnožení svým nositelům, ale proto, že umožnily rychlejší kladogenezi. Nelze tedy vyloučit, že nám náš organizmocentrický či alelocentrický pohled na evoluci dává falešné odpovědi, aniž by ovšem dosud publikované příklady druhového výběru byly skutečně průkazné. Vraťme se k antilopám: v okamžiku, kdy obě skupiny vznikly (předpokládejme, že jsou stejně staré), reprezentoval každou z nich jediný druh (tedy 1:1). Impaly se příliš nerozrůzňovaly, kdežto buvolci rozjeli výraznou speciaci po nějaké době tak dospějeme k poměru počtu druhů 1:5, potom 1:10, nebo třeba 2:25 a tak dále. Vykazují-li buvolci nějakou vlastnost, která je odlišuje od impal, třeba způsob výživy (buvolci jsou specializovaní spásači trávy, impaly nespecializovaní požírači všeho zeleného), nezbývá než prohlásit, že specializace na spásání trávy v evoluci antilop přibývá. Děje se tak mechanizmem druhového výběru; my ovšem budeme mít vždy podvědomý sklon ptát se, v čem je pro jednotlivou antilopu výhodnější spásat pouze trávu než žrát všechno, co najde; a zeptáme-li se takto, jistě i na nějakou výhodu omylem přijdeme.
Je docela pravděpodobné, že občas bude přirozený výběr jedinců v konfliktu s druhovým výběrem. Tak například velká hmotnost těla savců je obvykle spojena s nízkou populační hustotou a pomalou obměnou populace, čímž velikost těla ovlivňuje sklony druhu speciovat i vymírat. Pokud se z nějakého důvodu vyplatí být velký, třeba kvůli predátorům nebo z důvodů termoregulačních, individuální selekce bude preferovat zvětšení těla jedince; tím ovšem také může ovlivnit populační charakteristiky celého druhu, a to obvykle dosti nešťastně směrem k vyššímu riziku vymření. Otázka pak je, která tendence vyhraje, která úroveň selekce určí fylogenetický osud druhu a jeho příslušníků. Paradoxně se pak může stát, že některé druhy přežijí právě proto, že jejich jedinci nebyli příliš selekčně úspěšní.
Kdo ví, kolik znaků, na jejichž adaptivní výhodnost se ptáme, vzniklo vlastně jako vedlejší produkt druhové selekce, a správná odpověď na otázku, k čemu jsou tyto vlastnosti dobré, by tedy zněla, že urychlují speciaci. Co když i zvětšování lidského mozku probíhalo druhovým výběrem? Pokud by velkohlavé druhy speciovaly rychleji (třeba proto, ze velkohlaví lidoopi byli sexuálně vybíravější a měli větší tendenci vybírat si manželky v těsném sousedství), muselo by velkohlavých druhů lidoopů postupně přibývat ačkoli by nikde nedocházelo k přímé konkurenci velko a malohlavých jedinců, v níž by velkohlaví vítězili. Velký mozek by pak vlastně nebyl svému nositeli k ničemu, a pokud mu k něčemu byl (nejspíš asi k podvádění bližních), tato výhoda původně nebyla motorem jeho zvětšování. Dodejme pro jistotu, že tato hypotéza o druhové selekci jako motoru zvětšování lidského mozku je pouze divoká spekulace, jejímž smyslem je ilustrovat, jak málo vlastně víme.
Ke stažení
 Článek ve formátu PDF [234,07 kB]
Článek ve formátu PDF [234,07 kB]
O autorech
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
