










Přežívání populací v ostrůvkovitém prostředí
Od té doby, co Thomas Kuhn publikoval svou Strukturu vědeckých revolucí, je módní slovo paradigma, a to nejen mezi filozofy a sociology bádajícími o vědě, ale i mezi vědci samotnými. Snad v žádné vědní disciplíně se však nemluví o změně paradigmatu tak často jako v evoluční biologii a v ekologii. (Zatímco v evoluční biologii se občas uvádí možnost změny paradigmatu v budoucnu, v ekologii se naopak často zmiňuje změna paradigmatu v minulosti.) Datuje se přibližně do první poloviny osmdesátých let. Myslí se tím posun od důrazu myšlení na stabilitu, rovnováhu a homogenitu směrem k důrazu na proměnlivost a nestálost, nelinearitu a prostorovou i časovou heterogenitu.
Není to samozřejmě tak, že by si teprve začátkem osmdesátých let ekologové všimli, že na rozdíl od modelů, vycházejících z předpokladu stability a homogenity, je příroda mozaikovitá a proměnlivá. Spíš se zjistilo, že heterogenita a proměnlivost jsou něco zásadního a konstitutivního. Ta změna byla do značné míry určena i obyčejným technologickým pokrokem – před nástupem výpočetní techniky ekologické modely prostě musely být jednoduché, aby bylo možné je ověřovat. Ve hře ale asi byla i snaha opustit idealizující romantickou představu přírody jako harmonického celku, v němž jsou všechny procesy v rovnováze.
Posun paradigmatu se týkal především ekologie společenstev. Až do sedmdesátých let byla totiž společenstva považována za stabilní celky, jejichž jednotlivé části (populace) byly mezidruhovými vazbami více či méně těsně svázány. Ekologové se v té době tázali, čím je stabilita společenstev zajišťována a co umožňuje, že vedle sebe žijí různé druhy. Až později se začali tázat, zda jsou jednotlivé populace ve společenstvech tak pevně svázány a zda společenstva jsou vůbec v nějakém smyslu stabilní. Také populační biologové předpokládali, že populace jsou víceméně stabilní, pokud svou destruktivní činností nezasáhne člověk. Několik jevů ale svědčí o tom, že tento přístup je nedostatečný:
- Populace vznikají a zanikají, nejsou tedy úplně stabilní. Záleží samozřejmě na prostorovém měřítku a typu organizmu, nicméně lokální vyhynutí a opětovný vznik populace je pravidlem. Hmyzí populace na hektarové louce může po pár letech zmizet a za dalších několik let se zase objevit. U populací obratlovů ve stejně velkém lese to může trvat několik desítek let, nicméně stává se to. Nepřítomnost druhu na lokalitě nemusí svědčit o nevhodnosti prostředí, může být způsobena pouze náhodným vymřením.
- Řada lokálních populací se udržuje jen díky přistěhovalcům z okolí. Porodnost mnohdy bývá tak malá, že by k vyrovnání úmrtnosti nestačila a populace by za čas zanikla. Přítomnost druhu na lokalitě tedy nemusí svědčit o vhodnosti prostředí, ale o tom, že lokalita je blízko zdroje přistěhovalců.
- Dynamika populací probíhající ve velkých prostorových a časových měřítkách, v nichž hraje důležitou roli migrace. Když se například přemnoží hraboši, emigrují, a tím vzroste početnost hrabošů v sousedství. Vlna přemnožení se tak může šířit po krajině i stovky kilometrů daleko.
Podobná zjištění vedla ekology k zájmu o dynamiku populací v prostoru a v heterogenním prostředí. Časem vznikla vědní odvětví lišící se tím, na co kladou důraz. Například prostorová ekologie se zaměřuje na časoprostorovou dynamiku populací a předpokládá homogenní prostředí, v němž ovšem dynamika lokálních populací závisí na interakci se sousedními populacemi (viz zmíněné „migrační vlny“ hrabošů). Opačný přístup představuje krajinná ekologie (spíš její americká větev), která klade důraz na rozdílnou kvalitu různých typů prostředí, propojenost lokálních populací, typy rozhraní mezi jednotlivými prostředími a vůbec strukturu reálné krajiny. Největšího rozvoje ale asi doznala metapopulační ekologie. Ta má totiž nejen zásadní teoretické dopady, ale i jednoznačné praktické důsledky pro ochranu přírody.
Populace ve fragmentovaném prostředí
Metapopulační biologie se zabývá „sítí“ lokálních populací propojených migrací. Metapopulace je vlastně populace populací, respektive soubor lokálních populací propojených občasnou migrací. Migrace nemůže být nepřetržitá, poněvadž pak bychom měli jednu populaci, jejíž studium by bylo doménou zmíněné prostorové ekologie. Metapopulační ekologie tedy předpokládá, že místní populace budou odděleny, a studuje procesy probíhající na dvou úrovních – lokální a metapopulační. Nezajímá ji tvar a způsob propojení ostrůvků příznivého prostředí a většinou ani faktory týkající se dynamiky lokálních populací. Jde spíše o to, jak funguje celá metapopulace obývající fragmentované prostředí.Už nejjednodušší modely metapopulační dynamiky (viz rámeček 1 1 ) například ukázaly, že i když je dynamika lokálních populací náhodná, metapopulace může být stabilní. Při změně počtu osídlených ostrůvků má metapopulace tendenci vrátit se k původnímu stavu. Celé je to založeno na rovnováze kolonizace a vymírání. Intenzita kolonizace se zvýší, když se sníží počet obsazených ostrůvků, poněvadž tím se zvýší počet ostrůvků, které je možno kolonizovat. Ostrůvky, které nejsou obsazeny, slouží jako jakési rezervy pro případ vyhynutí na jiných ostrůvcích. Jestliže tedy chceme daný druh chránit, nelze chránit jen prostředí, kde se druh vyskytuje, ale je třeba chránit i prostředí, kde se nevyskytuje, avšak mohl by se tam uchýlit. Z modelů jen o trochu složitějších vyplynuly nelineární efekty, které provázejí zánik metapopulací i celých druhů.
Jak zaniká druh
Druh zanikne v okamžiku, když zaniknou všechny jeho populace. Jestliže je výskyt druhu omezen na malé území, stačí k jeho zániku drobná katastrofa. Občas ovšem vymizí druh z velkého území, a přitom si nevšimneme žádné změny prostředí, která by postihla všechny lokální populace najednou. Jak je to možné?Představme si, že se určitý druh vyskytuje v mnoha lokálních populacích. Občas nějaká lokální populace výrazně sníží početnost, nebo dokonce zmizí, ale díky migraci je znovu rychle dosycena. Závisí to na „zásobě migrujících“, a ta zas závisí na počtu lokálních populací. Když počet populací klesne pod určitou mez (která nemusí být nijak nízká), migrace mezi zbylými populacemi už nestačí náhodné početní propady v lokálních populacích dosycovat. Tak může zaniknout ještě více populací, čímž se ještě sníží počet migrantů, až celá metapopulace zanikne. Zánik metapopulace (a celého druhu) tedy může být spuštěn mírným snížením počtu osídlených ostrůvků nebo mírným snížením celkového počtu jedinců. Záleží na tom, kde je onen práh, za nímž už je metapopulace odsouzena k zániku.
Nevíme, jak často se tohle skutečně děje, ale lze tím dobře vysvětlit rychlé mizení nějakého druhu z oblasti bez viditelné příčiny. Podobné děje také pravděpodobně určují hranice areálů rozšíření jednotlivých druhů. Když se totiž podíváme na mapky areálů, všimneme si, že často „nedávají rozumný ekologický smysl“ – hranice areálů nekorespondují s nějakým zřejmým ekologickým rozhraním. Přítomnost druhu sama o sobě nemusí svědčit o vhodném prostředí – mnohé populace mohou přežívat v prostředí vlastně nepříznivém, syceném neustálým přílivem přistěhovalců. Naopak ani v příznivém prostředí daný druh nemusíme najít, pokud je toto prostředí příliš fragmentované a lokální populace jsou tak odsouzeny k postupnému vymizení. Vydejme se na hypotetickou cestu od středu areálu rozšíření nějakého druhu k okraji: Uprostřed areálu má druh většinou vysoké populační hustoty a vyskytuje se na mnoha lokalitách. Mezi nimi je neustálá migrace, takže nejde ani tak o metapopulaci jako o stabilní ostrůvkovitou populaci. Směrem k okraji areálu se populační hustoty snižují a intenzita migrace není tak velká, takže jednotlivé populace kolísají, občas i zaniknou. Mnohé přitom nejsou soběstačné, udržují se jen díky přílivu migrantů. Okraj areálu je pak vlastně hranicí, za níž příliv migrantů nestačí k dlohodobému udržení populací a četnost zániku lokálních populací není vyrovnávána četností vzniku nových. Snadno pochopíme, že hranice areálu v takovém případě nebude dána jen ekologickými podmínkami, ale hlavně vzdáleností od centra, schopností šíření a geografickými faktory, které šíření omezují, nebo podporují.
K čemu je dobrá fragmentace prostředí
Zatím to vypadalo tak, že s fragmentací prostředí jsou jen problémy – izolace malých populací vede k vymírání, které je horko těžko vyrovnáváno imigrací. Jak to ale v životě bývá, všechno zlé může být k něčemu dobré. Třeba v případě, že stejné prostředí obývají dva druhy, z nichž jeden ohrožuje druhý (ať už jde o dravce a jeho kořist či dva konkurenty, z nichž jeden je dominantní). Kdyby prostředí nebylo fragmentované, rychle by převládl ten „dominantní“. Ovšem v ostrůvkovitém prostředí by byla šance, že aspoň některé ostrůvky zůstanou neobsazené nepřítelem, a na nich lze přežívat. Pokud bude mít „dominantní“ druh nižší schopnost migrace, budou dlouhodobě oba druhy moci nerušeně koexistovat, poněvadž neosídlená místa budou představovat útočiště pro rychleji migrující druh.Tohle možná vypadá jako pouhá teoretická představa, ale ve skutečnosti je to zřejmě nejčastější způsob, jak mohou druhy žít společně. Jak totiž jinak vysvětlit to, že na mnoha lokalitách žijí druhy, o nichž víme, že jsou snadnou kořistí nějakého dravce nebo že mají nějakého silného konkurenta? Jednou možností je, že ten dravec či konkurent je sám nějak významně omezován (třeba dalším dravcem), druhou pak to, že se mu zkrátka ještě nepovedlo se na danou lokalitu dostat. A to je právě ono.
V praktické ochraně přírody se často zdůrazňuje nebezpečí izolace populací a společenstev. To je na jednu stranu oprávněné – skutečně izolované populace dřív nebo později vymřou. Populace mnoha druhů jsou ale izolovány pouze částečně, a dokonce díky této částečné izolaci druhy vůbec přežívají. Pokud fragmenty jejich prostředí propojíme koridory, můžeme jim zásadně uškodit, poněvadž podmínkou jejich existence je právě částečná oddělenost. Když si uvědomíme, jak velké ztráty biodiverzity jsou způsobeny invazemi (zhoubným pronikáním cizích druhů na území, kde se předtím nevyskytovaly), pochopíme nebezpečí umělého propojování.
- Co je horší – izolace, nebo propojenost? Záleží na dynamice druhů, které chceme chránit. Obecné řešení neexistuje. Pokud nějaké pravidlo vůbec platí, tak je to teze, že všechny změny jsou k horšímu. Když máme prostředí dlouhodobě fragmentované, je pravděpodobné, že druhy, které ho obývaly, jsou na fragmentaci adaptovány, nebo na ní dokonce závisejí. Druhy, kterým fragmentace prostředí vadila, tam už dávno nejsou. Propojení fragmentů tedy může jen uškodit. A jestliže naopak prostředí bylo původně souvislé, lze předpokládat, že fragmentace přinejmenším některým druhům bude vadit natolik, že ty druhy vymizí.
O složitosti
Metapopulační dynamika může být velmi složitá, navíc vychází z předpokladu, že i dynamika lokálních populací je složitá – přinejmenším není tak jednoduchá, že by vedla k rovnováze, naopak, vede k fluktuacím až k vymírání. Zároveň ovšem trochu paradoxně zajišťuje stabilitu celé metapopulace. Ta totiž může dlouhodobě přetrvávat, jen pokud dynamika jednotlivých lokálních populací není synchronizovaná. A to je zajištěno oddělením dynamiky jednotlivých populací díky fragmentaci prostředí a složitosti lokálních populačních dynamik. Takže složitost, která na jedné úrovni (lokální populace) vede k nestabilitě, na jiné stabilitu naopak zajišťuje. A ještě navíc díky fragmentaci prostředí, která je obecně považována za jasně negativní jev.Složitější modely metapopulační dynamiky ukazují ještě jednu důležitou věc. Pokud se týkají více navzájem na sebe působících druhů a obsahují nějaká omezení migrace (např. že se zvíře či rostlina dostane spíš na bližší než vzdálenější místo a že to nějakou dobu trvá), vedou k vzniku prostorové rozmanitosti a složitých postupně se proměňujících „obrazců“ vznikajících v ploše, a to i v případě, že na začátku mělo všechno stejnou strukturu. Jinými slovy ukazují, že sama různorodost prostředí může být důsledkem složité metapopulační dynamiky, a nikoli naopak. Složitost dynamiky v prostoru není něco, od čeho by šlo snadno odhlédnout, aniž bychom přestali rozumět tomu, co se v přírodě skutečně děje a může dít.
1. MODELOVÁNÍ METAPOPULAČNÍ DYNAMIKY
Na nejjednodušším modelu si můžeme ukázat základní principy ekologického modelování vůbec. Model se týká vztahu mezi dynamikou kolonizace, respektive vymírání, a počtem obsazených ostrůvků prostředí. Vychází z následujících předpokladů:
- Rychlost vymírání (tj. jak často nějaká lokální populace zanikne) závisí čistě na počtu už obsazených ostrůvků čím víc je jich obsazených, tím víc populací může zase zmizet. Rychlost vymírání je rovna E*P, kde E je druhově specifická pravděpodobnost vymření jedné populace a P je procento obsazených ostrůvků. Vztah mezi počtem obsazených ostrůvků a rychlostí vymírání je tedy lineární.
- Rychlost kolonizace je úměrná podílu už obsazených ostrůvků P (poněvadž s počtem populací roste počet možných kolonizátorů) a zároveň podílu ještě neobsazených ostrůvků 1-P, poněvadž čím více ostrůvků je neobsazených, tím více jich lze obsadit. Takže to můžeme vyjádřit jako K*P*(1-P), kde K je opět druhově specifická pravděpodobnost obsazení nového ostrůvku. Vztah mezi rychlostí kolonizace a počtem obsazených ostrůvků je parabola obrácená směrem dolů, s maximem na polovině obsazených ostrůvků.
- >
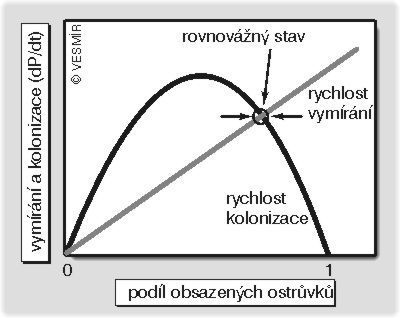
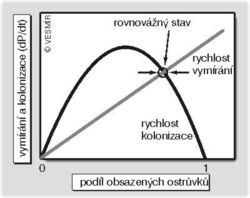 Celková změna počtu obsazených ostrůvků je rovna rozdílu mezi rychlostí vymírání a rychlostí kolonizace. Můžeme to napsat jako diferenciální rovnici dP/dt = K*P*(1 p) E*P. Bod, kde se protíná křivka vyjadřující rychlost kolonizace s křivkou vyjadřující rychlost vymírání, představuje rovnovážný stav, tj. odpovídá tomu relativnímu počtu obsazených ostrůvků, na němž se metapopulační dynamika ustálí (pokud by ale pravděpodobnost vymření jedné populace byla větší než pravděpodobnost obsazení nového ostrůvku, tímto bodem by byla jen nula, tj. celá metapopulace by vymírala a vymřela). Jestliže se náhodou tento počet zvýší, klesne rychlost kolonizace a stoupne rychlost vymírání, a tím se počet opět sníží (a naopak). Z modelu je jasné, že kdyby byly obsazeny všechny ostrůvky, nešlo by o rovnovážný stav, poněvadž pak by bylo možné pouze vymírání a nikoli kolonizace. Umělá likvidace ostrůvků (i neobsazených!) vede k posunu rovnovážného bodu směrem k menšímu počtu obsazených ostrůvků (než se ustálí nový rovnovážný stav, může docházet jen k vymírání).
Celková změna počtu obsazených ostrůvků je rovna rozdílu mezi rychlostí vymírání a rychlostí kolonizace. Můžeme to napsat jako diferenciální rovnici dP/dt = K*P*(1 p) E*P. Bod, kde se protíná křivka vyjadřující rychlost kolonizace s křivkou vyjadřující rychlost vymírání, představuje rovnovážný stav, tj. odpovídá tomu relativnímu počtu obsazených ostrůvků, na němž se metapopulační dynamika ustálí (pokud by ale pravděpodobnost vymření jedné populace byla větší než pravděpodobnost obsazení nového ostrůvku, tímto bodem by byla jen nula, tj. celá metapopulace by vymírala a vymřela). Jestliže se náhodou tento počet zvýší, klesne rychlost kolonizace a stoupne rychlost vymírání, a tím se počet opět sníží (a naopak). Z modelu je jasné, že kdyby byly obsazeny všechny ostrůvky, nešlo by o rovnovážný stav, poněvadž pak by bylo možné pouze vymírání a nikoli kolonizace. Umělá likvidace ostrůvků (i neobsazených!) vede k posunu rovnovážného bodu směrem k menšímu počtu obsazených ostrůvků (než se ustálí nový rovnovážný stav, může docházet jen k vymírání).
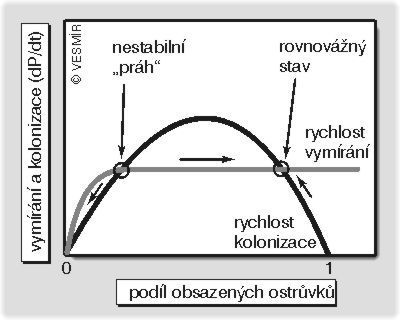
 Tenhle model lze všelijak zesložiťovat. Můžeme si třeba představit, že rychlost vymírání neporoste s počtem obsazených ostrůvků lineárně, poněvadž do hry vstoupí pozitivní zpětná vazba mezi počtem obsazených ostrůvků a migrací. Při dostatečném počtu obsazených ostrůvků bude celková intenzita migrace už tak velká, že i kdyby nějaká lokální populace spěla k zániku, příliv přistěhovalců tomu zabrání. Potom se může stát, že se obě křivky protnou vícekrát. Pak budou existovat dva stavy, kdy je kolonizace a vymírání v rovnováze, ovšem ten první je nerovnovážný: při zvýšení počtu obsazených ostrůvků převáží intenzita kolonizace nad intenzitou vymírání, a tak se počet obsazených ostrůvků bude zvyšovat až k druhé rovnováze. Naopak i velmi malé snížení počtu obsazených ostrůvků pod onen nestabilní práh povede k vymizení celé populace, poněvadž intenzita vymírání již nebude vyrovnána kolonizací. To je ta nelinearita. Anebo by třeba populace nemusela úplně vymizet, ale daný druh by přežíval na nějakých zvláště příznivých ostrůvcích. Kdybychom tenhle efekt útočišť (refugií) chtěli vyjádřit v modelu, mohli bychom třeba předpokládat ještě složitější křivku intenzity vymírání, která by zpočátku téměř nestoupala, poněvadž by v této části zobrazovala místa, kde k vymírání téměř nedochází.
Tenhle model lze všelijak zesložiťovat. Můžeme si třeba představit, že rychlost vymírání neporoste s počtem obsazených ostrůvků lineárně, poněvadž do hry vstoupí pozitivní zpětná vazba mezi počtem obsazených ostrůvků a migrací. Při dostatečném počtu obsazených ostrůvků bude celková intenzita migrace už tak velká, že i kdyby nějaká lokální populace spěla k zániku, příliv přistěhovalců tomu zabrání. Potom se může stát, že se obě křivky protnou vícekrát. Pak budou existovat dva stavy, kdy je kolonizace a vymírání v rovnováze, ovšem ten první je nerovnovážný: při zvýšení počtu obsazených ostrůvků převáží intenzita kolonizace nad intenzitou vymírání, a tak se počet obsazených ostrůvků bude zvyšovat až k druhé rovnováze. Naopak i velmi malé snížení počtu obsazených ostrůvků pod onen nestabilní práh povede k vymizení celé populace, poněvadž intenzita vymírání již nebude vyrovnána kolonizací. To je ta nelinearita. Anebo by třeba populace nemusela úplně vymizet, ale daný druh by přežíval na nějakých zvláště příznivých ostrůvcích. Kdybychom tenhle efekt útočišť (refugií) chtěli vyjádřit v modelu, mohli bychom třeba předpokládat ještě složitější křivku intenzity vymírání, která by zpočátku téměř nestoupala, poněvadž by v této části zobrazovala místa, kde k vymírání téměř nedochází.
 Klasická (levinsovská) metapopulace předpokládá náhodnou dynamiku lokálních populací, což by dávalo obrázek podobný nejstaršímu nortonovskému šetřiči obrazovky populace (představované hvězdičkami) se náhodně objevují a náhodně mizejí, přesto jich je pořád přibližně stejně a jsou víceméně rovnoměrně rozptýleny. Ovšem model předpokládající nestejnost jednotlivých ostrůvků by zahrnoval dlouhodobé přetrvávání v nejkvalitnějších ostrůvcích, šíření do těch méně kvalitních, opětovný ústup atd. Složitější modely metapopulační dynamiky (už nikoli analytické, ale spíše simulační) zahrnují ještě roli odlišné vzdálenosti (to znamená izolovanosti) jednotlivých ostrůvků, vnitřní dynamiku lokálních populací, určité rozložení vzdáleností, kam až může jedinec domigrovat atd.
Klasická (levinsovská) metapopulace předpokládá náhodnou dynamiku lokálních populací, což by dávalo obrázek podobný nejstaršímu nortonovskému šetřiči obrazovky populace (představované hvězdičkami) se náhodně objevují a náhodně mizejí, přesto jich je pořád přibližně stejně a jsou víceméně rovnoměrně rozptýleny. Ovšem model předpokládající nestejnost jednotlivých ostrůvků by zahrnoval dlouhodobé přetrvávání v nejkvalitnějších ostrůvcích, šíření do těch méně kvalitních, opětovný ústup atd. Složitější modely metapopulační dynamiky (už nikoli analytické, ale spíše simulační) zahrnují ještě roli odlišné vzdálenosti (to znamená izolovanosti) jednotlivých ostrůvků, vnitřní dynamiku lokálních populací, určité rozložení vzdáleností, kam až může jedinec domigrovat atd.
2. HISTORICKÉ ZDROJE METAPOPULAČNÍ EKOLOGIE
I když jde o obor rozvíjející se hlavně v 90. letech 20. století, už v 50. letech upozornili australští ekologové H. G. Andrewartha a L. C. Birch na skutečnost, že lokální hmyzí populace jsou často efemérní (pomíjivé) a kolísání jejich početnosti nesvědčí o hustotně závislé regulaci, ale spíš odráží vlivy počasí. To, že nedochází ke globálnímu vymírání, vysvětlovali tím, že jedna populace zanikne, jiná zas vznikne. Neformulovali to matematicky, ale jejich představa v podstatě odpovídala klasickému modelu metapopulace.
- Rovnovážná teorie ostrovní biogeografie, jejímiž autory jsou Robert MacArthur a Edward O. Wilson, upozornila r. 1963 nejen na to, že populace vskutku vymírají, ale hlavně na to, že znalost dynamiky vymírání a kolonizace je důležitá pro pochopení některých pozoruhodných jevů v živé přírodě. Vysvětluje totiž počet druhů na ostrovech rovnováhou mezi kolonizací a vymíráním, která závisí na velikosti ostrova a jeho vzdálenosti od pevniny (čím menší ostrov, tím rychlejší vymírání a čím vzdálenější ostrov, tím pomalejší kolonizace).
- Levinsův model metapopulační dynamiky je též založen na rovnováze mezi kolonizací a vymíráním. Na rozdíl od ostrovní teorie ale zůstal po léta téměř nepovšimnut.
SLOVNÍČEK
společenstvo - soubor populací různých druhů žijících společně na jednom místě
populace - skupina jedinců téhož druhu žijící v téže době na tomtéž místě
metapopulace - soubor oddělených populací téhož druhu, mezi nimiž probíhá migrace
Ke stažení
 Článek ve formátu PDF [117,63 kB]
Článek ve formátu PDF [117,63 kB]
O autorovi
David Storch

Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
