










O vzniku druhů přirozeným výběrem
Vznik druhů byl donedávna považován především za vedlejší produkt geografické izolace populací. Dnes se zdá, že překvapivě velkou roli může hrát prostředí, přesně řečeno rozmanitost způsobů, jak může daný druh prostředí využívat. Poznatek, že druhy často vznikají aktivním přizpůsobením rozdílným typům prostředí bez geografické izolace, patří k nejpřevratnějším zjištěním evoluční biologie a ekologie poslední dekády 20. století.
Každého, kdo čte monumentální Darwinovo dílo, ohromí jednak autorův neuvěřitelný pozorovací talent, jednak hloubka vhledu do přírodních procesů – v ekologii je jen málo jevů, kterých si Darwin nevšiml, a málo procesů, jejichž potenciální důležitost nezmínil. Čtenáře ale možná poněkud překvapí, jak malou pozornost věnoval Darwin vlastnímu procesu vzniku druhů. Darwinovo dílo se skutečně vzniku druhů, jak jej dnes chápeme, téměř vůbec netýká – Darwin popisoval (a vysvětloval) postupné změny v rámci jedné evoluční linie, zatímco štěpení evolučních linií (viz rámeček „Anageneze…“) stálo stranou jeho zájmu. Předpokládal totiž, že pokud evoluce spočívá v přizpůsobování prostředí, tak vlastní vznik druhů, speciace, nepředstavuje vůbec žádný problém – část jedinců se zkrátka někdy přizpůsobí jednomu typu prostředí, část jinému, a tak vzniknou dva druhy. Darwin totiž nebral moc vážně existenci druhů a mezi vnitrodruhovou a mezidruhovou proměnlivostí viděl kontinuum.
Dnes víme, že tak jednoduché to není. Druhy jsou přecejen něco víc než skupiny vzájemně si podobných jedinců, kteří mají společný původ. Přinejmenším pohlavně se rozmnožující jedinci si mezi sebou v rámci druhu neustále vyměňují genetickou informaci, geneticky (i jinak) spolu komunikují. A při vzniku druhů (tedy rozštěpení původního druhu na dva nebo více dceřiných druhů) jde právě o zpřetrhání této komunikace. Problém je jak přinutit jedince právě vznikajících druhů, aby se spolu přestali křížit. Ten problém je tak velký (viz rámeček „Křížení…“), že donedávna se za jediný opravdu fungující způsob jak zajistit reprodukční izolaci nově vzniklých druhů považovala fyzická izolace populací. A tak se za hlavní způsob vzniku druhů pokládala speciace založená na dostatečně dlouhé geografické izolaci populací (viz rámeček „Speciace“).
Klasická představa vzniku druhů
Podle klasické představy jsou nejprve dvě nebo více populací vlivem vnějších podmínek izolovány geograficky – např. se oddálí kontinenty, které byly původně spojeny, vyvrásní se pohoří nebo se rozpadne souvislý areál druhu. Každá populace se pak vyvíjí odděleně a trochu jinak – ať už proto, že se přizpůsobuje jinému prostředí, nebo proto, že ji ovlivňují náhodné genetické procesy. Časem se populace rozrůzní natolik, že přestanou být vzájemně slučitelné – buď už se spolu jedinci z oněch oddělených populací vůbec nemohou křížit (tak, aby měli plodné potomstvo), anebo se sice křížit mohou, ale jejich potomci už jsou příliš znevýhodněni. Ti pak mají málo potomků, takže přirozený výběr časem vede k zamezení mezipopulačního (a teď už mezidruhového) křížení (s tím je ale trochu problém, viz rámeček „Křížení…“).Tato představa vzniku druhů nebyla založena jen na nějakých teoretických genetických úvahách, ale hlavně na empirických znalostech geografického rozšíření rostlin a živočichů. Dlouho se totiž vědělo, že na izolovaných plochách, jako jsou ostrovy, žijí jiné druhy než na pevnině (Vesmír 77, 362, 1998/7). Na vzdálenějších ostrovech je přitom více endemitů (tj. druhů, které se nikde jinde nevyskytují) než na bližších, a navíc ti endemiti vznikají přednostně ve skupinách, které se hůře šíří. Izolovanost tedy evidentně vede ke vzniku druhů. Na základě podobných pozorování Ernst Mayr, jeden z tvůrců nové neodarwinistické evoluční syntézy, prosazoval myšlenku, že geografická izolace je nejdůležitějším faktorem ovlivňujícím vznik druhů. Mayr přitom zavedl a prosazoval především peripatrický model speciace (viz rámeček „Speciace“), kdy nové druhy vznikají z malých izolovaných populací na okraji areálu rozšíření původního druhu.
Všimněme si, že tato klasická představa speciace vlastně považuje vznik druhů za cosi, co se děje víceméně omylem, vinou vnějších okolností – speciace je vedlejší produkt dlouhotrvajícího oddělení populací. V žádném případě druhy podle této představy nevznikají „na objednávku prostředí“, zaplňováním předem daných ekologických nik. To má poměrně dalekosáhlé ekologické důsledky. Ekologické niky podle tohoto modelu neexistují předem, ale vznikají až interakcí jednotlivých druhů s prostředím – druhy, které shodou okolností vznikly, si mezi sebou prostředí nějak rozdělují, a toto rozdělení může být do značné míry náhodné.
Může přirozený výběr vést ke speciaci?
Během posledních deseti let se zjistilo, že to tak jednoznačné být nemusí. Našly se totiž druhy, které mají dvě nebo více forem, z nichž každá se specializuje trochu na něco jiného a podle toho se také odlišuje. Křivka Loxia curvirostra například v Americe vytváří několik forem, které se liší tvarem zobáku a specializují se na šišky různých druhů stromů (Vesmír 79, 486, 2000/9). Africký pěvec bulbul malý (Andropadus virens) má zase dvě formy – jedna žije v pralese a druhá v přechodové zóně mezi pralesem a savanou – a liší se velikostí i morfologicky (viz Vesmír 77, 356, 1998/6). Něco podobného lze pozorovat u australského scinka Carlia rubrigularis, u něhož se přišlo na to, že ekologické podmínky určují vzhled scinků mnohem výrazněji než geografická izolace – což je přesný opak Mayrova pozorování. V obou posledně zmíněných případech se také zjistilo, že dvě ekologicky odlišné formy se udržují, přestože mezi nimi existuje tok genů – ty formy nejsou reprodukčně oddělené. To je velmi důležité, poněvadž to znamená, že odlišné selekční tlaky stačí k tomu, aby se populace rozdělila na různé ekologické formy, i když se jejich genetický materiál pořád trochu míchá. A zároveň to znamená, že když prostředí umožňuje více typů adaptací, které se vzájemně vylučují, tak se tyto možnosti někdy mohou realizovat. Jinými slovy když je v prostředí více odlišných ekologických nik, které druh může využívat, někdy je skutečně využívat začne.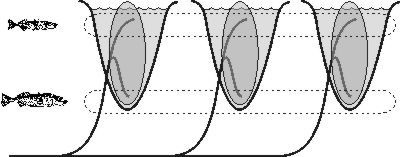{kind=link}
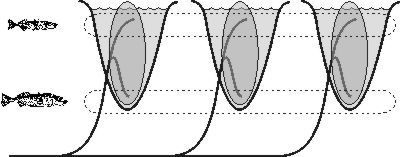
Tyto případy sice znamenají, že prostředí může vést k částečnému ekologickému rozdělení původního druhu, ale nijak z nich neplyne, že dojde ke speciaci, tedy k úplnému reprodukčnímu oddělení. K tomu je totiž ještě třeba zařídit, aby se obě formy mezi sebou přestaly křížit neboli aby jedinci měli tendenci přednostně se křížit s jedinci náležícími k téže formě. To se ovšem u těch druhů, kde existuje pohlavní výběr, stává často. Někdy se pohlavní výběr týká přímo těch znaků, které souvisejí se získáváním zdrojů, a pak je to jednoduché – jakmile se vytvoří dvě formy, jež se liší ve způsobech získávání zdrojů, automaticky se mezi sebou méně kříží. Pohlavní výběr je zcela klíčový; dokonce se má za to, že je mnohem důležitější než ekologická specializace – druhy mohou divergovat i bez ní, když je u nich silná tendence vybírat partnera, který vypadá podobně jako vybírající si jedinec (viz Vesmír 79, 366, 2000/7).
Asi nejhezčí a nejlépe dokumentovaný případ ekologické sympatrické speciace představují koljušky, které po skončení poslední doby ledové osídlily sladkovodní jezera v kanadské Britské Kolumbii. Z původně mořské koljušky tříostné (Gasterosteus aculeatus) vznikly v každém jezeru dvě formy lišící se velikostí: větší osídluje dno a živí se bentickými živočichy (obývajícími bahnité dno), menší se pohybuje ve vodním sloupci a živí se planktonem. Koljušky žijící v jednom jezeru jsou si přitom navzájem příbuznější než koljušky stejné formy žijící v různých jezerech, takže se dá předpokládat, že alespoň v některých jezerech vznikly obě formy sympatricky. Tyto dvě formy se přitom navzájem téměř nekříží – dávají přednost partnerům náležícím ke stejné formě – a když je zkřížíme uměle, hybridi jsou značně znevýhodněni, poněvadž nejsou schopni dobře se živit planktonem ani bentickými živočichy. Zvláště kuriózní je skutečnost, že geneticky nepříbuzní příslušníci téže formy z různých jezer se kříží a rozmnožují zcela bez problémů. Kdybychom tedy na ně aplikovali klasickou definici biologického druhu (jejímž autorem je mimochodem právě zmíněný Ernst Mayr), budou jedinci dané formy z různých jezer tvořit jeden druh, přestože jsou si vzájemně méně příbuzní než jedinci různých forem žijící v jednom jezeru (obr. 1).
{kind=link}
Zdá se tedy, že přirozený výběr, který zvýhodňuje jedince různým způsobem přizpůsobené prostředí, může vést ke vzniku druhů. Podmínkou je, aby se různé typy přizpůsobení vzájemně vylučovaly (jako třeba ty, které souvisejí s různou tělesnou velikostí), poněvadž jinak by převážil typ schopný využívat všechno. Otázkou ovšem zůstává, jak často takhle druhy skutečně vznikají.
Jak zjišťovat, co se vlastně v evoluci stalo
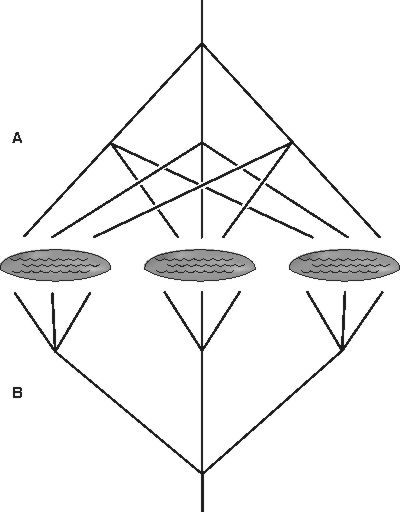
Při rekonstrukci evoluce jsme často odkázáni na hodně nepřímé doklady. Pro vznik druhů máme k dispozici vlastně jenom jejich současné rozšíření, někdy nanejvýš doplněné fosilními nálezy. Současné rozšíření přitom může celkem dobře svědčit ve prospěch alopatrické speciace (když se příbuzné druhy vyskytují v úplně jiných oblastech), sympatrickou speciaci lze ale doložit hůře, poněvadž samotný společný výskyt druhů už může být druhotný. Dobrým dokladem sympatrické speciace je jen ten případ, kdy se skupina blízce příbuzných druhů nachází v jednom místě, o němž lze stěží předpokládat, že by tam populace mohly být geograficky izolovány. Cichlidy žijící v afrických kráterových jezerech jsou si například blízce příbuzné v rámci jezera, ale méně příbuzné mezi jezery (při alopatrické speciaci by tomu mělo být naopak; (obr. 2). Obecně by alopatrická speciace měla vést k tomu, že bližší příbuzní budou více geograficky odděleni než vzdálení příbuzní.Pro význam sympatrické speciace při vzniku druhů může svědčit i skutečnost, že skupiny, u nichž existuje pohlavní výběr, mají často větší druhové bohatství než sesterské skupiny. Třeba u ptáků se zjistilo, že skupiny s výrazným pohlavním dimorfizmem (tj. kde je sameček barevnější než samička), u nichž se předpokládá pohlavní výběr, jsou skutečně druhově bohatší. Druhové bohatství cichlid ve Viktoriině jezeře se také vysvětluje jejich tendencí k výběru stejně zbarveného partnera (a jejich barevnou variabilitou, viz Vesmír 78, 338, 1999/6). Pohlavní výběr tedy asi opravdu vede ke vzniku druhů.
Co z toho plyne
Zdá se, že sympatrická speciace je v přírodě mnohem běžnější, než se ještě nedávno myslelo. Její relativní význam ve srovnání s jinými typy speciace lze sice těžko posoudit, může však být značný. Důležité je ale hlavně to, že prostředí může hrát roli při vzniku druhů, že druhy nemusejí vznikat jen jako náhodný produkt geografické izolace, ale mohou aktivně vyplňovat alternativní možnosti, které jim prostředí nabízí. Evoluce druhového bohatství tedy může velmi těsně souviset s různorodostí prostředí, tak jak je vnímána jednotlivými druhy. Vznik druhů může být proces nejen aktivní, ale, jak dokládá příklad koljušek, do jisté míry i zákonitý.Anageneze a kladogeneze
Evoluce živých forem má dva aspekty. Jedním je postupná změna (morfologická, fyziologická, etologická) v rámci jedné evoluční linie. Tomuto procesu říkáme anageneze a právě tou se zabýval Darwin. Štěpení evolučních linií, tedy větvení „evolučního stromu“, pak nazýváme kladogeneze. Tu primárně způsobuje rozpad jednoho druhu na více druhů dceřiných, který nemusí souviset s přirozeným výběrem a přizpůsobováním prostředí. Jestliže se anageneze a kladogeneze nerozlišují, vede to k neporozumění. Darwinovi se například vyčítá, že jeho teorie nepředpovídá žádný konkrétní nárůst diverzity, přesně řečeno vůbec žádné jevy související s diverzitou. To je sice pravda, ale je to zkrátka tím, že Darwinova teorie se vznikem diverzity vůbec nezabývá.
Křížení, fitness a vazby mezi geny
Na první pohled je to jednoduché. Mezidruhoví kříženci jsou nějakým způsobem znevýhodněni, mají méně potomků, a tak přirozený výběr působí proti mezidruhovému křížení. Takhle jednoduše to ale nefunguje proto, že jednotkou přirozeného výběru ve skutečnosti nejsou jedinci (ti nakonec umřou všichni), ale jejich geny, přesněji řečeno alely (různé verze genů). A ty jsou (jak praví třetí Mendelův zákon) volně kombinovatelné. K tomu, aby se dva právě vznikající druhy přestaly křížit, je třeba – poněkud zjednodušeně řečeno – odstranit z populace alely (vlohy), které je k mezidruhovému křížení předurčují. Jenže to se právě nepodaří, pokud budou mezidruhoví kříženci znevýhodněni jen mírně – to, že budou mít méně potomků, nezaručí, že všechny tyto alely z populace vymizí. Ony totiž vymizí alely, které (negativně) ovlivňují fitness hybridů, ale ty, co ovlivňují mezidruhové křížení, se dál budou udržovat v populaci právě proto, že jsou na nich nezávislé. Odstraní se z populace jedině tehdy, pokud budou s alelami ovlivňujícími fitness ve vazbě, tj. budou třeba na tomtéž chromozomu. To je ale do značné míry věc náhody.
Když se začalo ukazovat (viz hlavní text), že sympatrická speciace je zřejmě mnohem běžnější, než se zdálo, začal se hledat model, který by to vysvětloval z genetického hlediska. A jak to tak bývá, našel se rychle. Je to jednoduché – celé to bude fungovat, pokud bude více genů ovlivňujících fitness, a méně (nejlépe jeden) těch, které ovlivňují mezidruhové křížení. Pak je totiž velká pravděpodobnost, že alely ovlivňující negativně mezidruhové křížení se dostanou do vazby aspoň s některou alelou negativně ovlivňující fitness hybridů, a tak se s nimi „svezou“.
Speciace
Speciace může být buď alopatrická, nebo sympatrická. Alopatrická znamená, že nové druhy vznikají v různých oblastech, zatímco sympatricky vznikají druhy bez předchozí geografické izolace. Do alopatrické speciace patří také speciace peripatrická, kdy druhy vznikají z malých izolovaných populací na okraji areálu rozšíření (zde se uvažuje o významné roli genetického driftu, díky němuž se genetická struktura malé populace rychle – a náhodně – mění), odkud se později rozšíří na území mateřského druhu. Jiným typem alopatrické speciace je speciace extinkční, kdy druh na většině svého areálu rozšíření vymře a nové druhy postupně vzniknou z izolovaných zbytků původního rozšíření.
Sympatrická speciace může být způsobena zkřížením dvou druhů, hybridizací, a vznikem životaschopného tvora, který už se není schopen úspěšně křížit s žádným z otcovských druhů. To často souvisí s polyploidizací, tedy zmnožením chromozomů, které právě vede k nekompatibilitě s otcovskými druhy. Podobných typů speciace je více. Z našeho hlediska je ovšem důležitější tzv. ekologická nebo kompetiční speciace, kdy uvnitř druhu nejprve probíhá rozdílná ekologická specializace, a ta omezí vnitrodruhovou konkurenci.
Ke stažení
 Článek ve formátu PDF [135,12 kB]
Článek ve formátu PDF [135,12 kB]
O autorovi
David Storch

Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
