










Embryologie a genetika
V roce 1822 známý francouzský přírodovědec E. Geoffroy Saint-Hilaire uveřejnil v časopise Mémoires de la Muséum de Histoire Naturelle obsáhlé pojednání o tom, že ventrální (břišní) strana těla členovců odpovídá hřbetní (dorzální) straně těla obratlovců a naopak. Na anatomii humra (obr. obrázek) ukázal, že v obrácené poloze je jeho nervová soustava nahoře, nad zažívací trubicí, a pod tou je srdce, podobně jako je tomu např. u savců. Tento názor o dorzoventrální inverzi tělní organizace mezi členovci a obratlovci podporoval jeho koncepci jednotné základní stavby všech mnohobuněčných živočichů (Metazoa). Jeho článek vyvolal ve francouzské Akademii polemiku s proslulým G. Cuvierem. 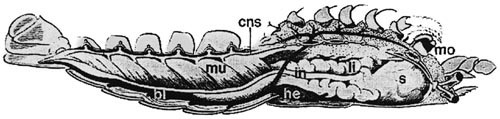
{kind=link}
{kind=link}
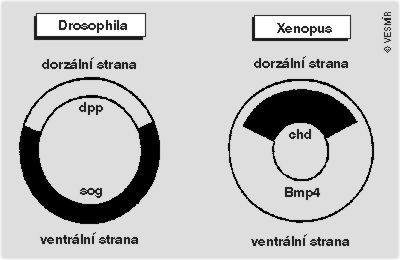
Antagonizmu mezi faktory Sog a Dpp u drozofily odpovídá u obratlovců antagonizmus mezi Chd a Bmp-4. Vzdor morfologickým rozdílům mezi členovci a obratlovci funkční chování genů sog/chd a dpp/bmp-4 naznačuje homologní vývojové mechanizmy při vzniku dorzoventrality, které musely existovat již u společného předka, z něhož obě skupiny na počátku kambria vznikly. Ačkoli neurogeneze u hmyzu a neurální indukce u obratlovců byly pokládány za odlišné procesy, na molekulární úrovni jsou zřejmě v obou případech řízeny shodným evolučně konzervovaným systémem. K obrácení dorzoventrální osy v evoluci došlo v gastrulaci a vedlo k oddělení dvou hlavních skupin mnohobuněčných živočichů – prvoústých (Prostomia) a druhoústých (Deuterostomia). U prvně jmenovaných se prvoústa gastruly stávají ústy živočicha, jak je tomu u většiny kmenů bezobratlých. U druhoústých se prvoústa gastruly stávají řitním otvorem živočicha a jeho ústa vznikají druhotně na opačném konci těla. Tato situace je typická pro obratlovce, ostnokožce a několik menších kmenů bezobratlých (viz článek Antona Markoše na s. 89). K oddělení prvoústých a druhoústých došlo asi před 565 až 530 miliony let.
Jiný příklad homologického procesu na molekulární úrovni je vývoj složeného oka hmyzu a komorového oka obratlovců. Doposud byly oba procesy pokládány za konvergenci, jak se ale ukázalo, jsou řízeny konzervovaným genem Pax-6/eyeless (Vesmír 74, 495, 1995/9). V obou případech vznikly oči z fotoreceptorů společného předka. E. M. De Robertis a Y. Sakai navrhují pro hypotetického předka prvoústých a druhoústých označení Urbilateria (primitivní bilaterální živočich). Vyznačuje se těmito společnými znaky:- jeho předozadní polarita je určována komplexy homeoboxových genů (Hox), s kterými spolupracují další geny,
- má konzervovaný systém určující dorzoventrální uspořádání antagonistickými produkty genů sog/chd a dpp/bmp-4 jako mimobuněčnými signály,
- má podpokožkový podélný centrální nervový systém,
- funkce genů Pax-6/eyeless a přítomnost proteinů opsinů naznačuje, že ze světločivných buněk tohoto předka se vyvinulo jak složené oko hmyzu, tak komorové oko obratlovců,
- měl též oběhový systém s kontraktilními cévami, jejichž vývoj je řízen homeoboxovými geny, homologickými u členovců i obratlovců.
Pro posouzení vztahů a rozlišení různých živočišných kmenů v průběhu evoluce je pozoruhodným přínosem porovnání sekvencí bází menší podjednotky ribozomální ribonukleové kyseliny (18S rRNA) u mnohobuněčných živočichů (obr. obrázek).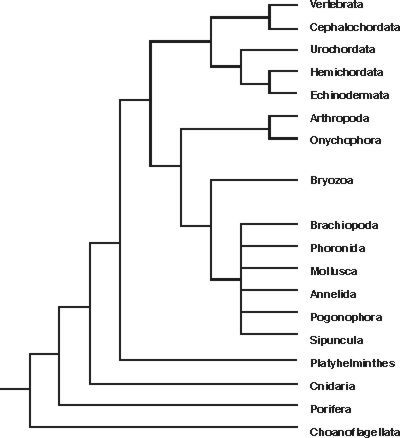{kind=link}
 Rozdíly sekvencí umožňují posoudit větvení kmenů. Mnohobuněční živočichové (Metazoa) vznikli asi před jednou miliardou let oddělením od skupiny Choanoflagelata a při tzv. kambrické explozi (zhruba před 540 miliony lety) došlo k rozlišení živočišných kmenů a ty v podstatě zůstaly uchovány až do současnosti. Pozoruhodné také je, že od té doby se nový živočišný kmen neobjevil – řídící systémy znamenaly zřejmě určitá vývojová omezení pro evoluci. Snad u všech mnohobuněčných určují maternální a časné embryonální řídící geny tělní osy i hlavní rysy uspořádání těla. Mnoho z nich obsahuje charakteristické sekvence bází – homeoboxy (Vesmír 74, 276, 1995/5). Tyto geny aktivují nekolik tříd vysoce konzervovaných genů, tzv. homeoboxových (Hox) genů. Určují identitu různých oblastí těla (u členovců např. segmentů nesoucích končetiny, popř. křídla, tykadla apod.). Na základě určení polohy v těle se pak aktivují specifické cílové geny. Shluky Hox-genů mají u různých živočišných kmenů shodné pořadí v chromozomech a jsou aktivovány popořadě, ve shodě např. s předozadní osou těla. Jejich dnešní řídící úloha nemusí však vždy odpovídat jejich původní úloze u předků. Např. ve vývoji končetin obratlovců se uplatňují nejen geny příbuzné genům drozofily, ale i genům hlístice Caenorhabditis elegans, která nohy nemá. Jejich evoluční počátek předchází vzniku končetin.
Rozdíly sekvencí umožňují posoudit větvení kmenů. Mnohobuněční živočichové (Metazoa) vznikli asi před jednou miliardou let oddělením od skupiny Choanoflagelata a při tzv. kambrické explozi (zhruba před 540 miliony lety) došlo k rozlišení živočišných kmenů a ty v podstatě zůstaly uchovány až do současnosti. Pozoruhodné také je, že od té doby se nový živočišný kmen neobjevil – řídící systémy znamenaly zřejmě určitá vývojová omezení pro evoluci. Snad u všech mnohobuněčných určují maternální a časné embryonální řídící geny tělní osy i hlavní rysy uspořádání těla. Mnoho z nich obsahuje charakteristické sekvence bází – homeoboxy (Vesmír 74, 276, 1995/5). Tyto geny aktivují nekolik tříd vysoce konzervovaných genů, tzv. homeoboxových (Hox) genů. Určují identitu různých oblastí těla (u členovců např. segmentů nesoucích končetiny, popř. křídla, tykadla apod.). Na základě určení polohy v těle se pak aktivují specifické cílové geny. Shluky Hox-genů mají u různých živočišných kmenů shodné pořadí v chromozomech a jsou aktivovány popořadě, ve shodě např. s předozadní osou těla. Jejich dnešní řídící úloha nemusí však vždy odpovídat jejich původní úloze u předků. Např. ve vývoji končetin obratlovců se uplatňují nejen geny příbuzné genům drozofily, ale i genům hlístice Caenorhabditis elegans, která nohy nemá. Jejich evoluční počátek předchází vzniku končetin.
U ploštěnců (Plathelminthes) obsahuje shluk Hox jen 4 geny, u drozofily je jich 8, u obratlovců 38 ve 4 různých shlucích. Společný předek prvoústých a druhoústých před kambrickou explozí měl asi shluk o nejméně 6 genech Hox. V různých liniích došlo k zdvojení genů Hox v souvislosti se vzrůstající složitostí tělesné organizace – u obratlovců se zmnožily celé shluky (obr. obrázek). Homeotické geny byly nalezeny u všech živočišných kmenů s výjimkou hub (Porifera). 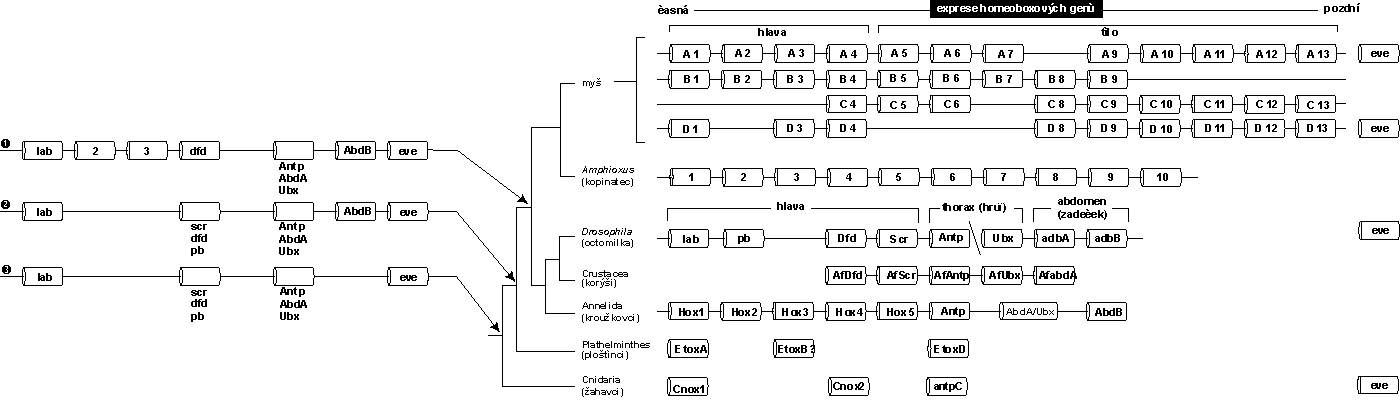
{kind=link}
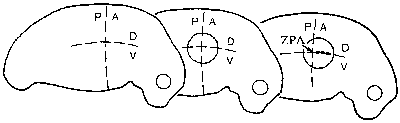
Morfogenetická pole neboli oblasti v tkáni s koncentračním spádem produktu některého regulačního genu se těšily velkému zájmu experimentální embryologie. Teprve molekulární embryologie jim však dala nový rozměr na základě časově a prostorově řízeného projevu různých genů a šíření jejich produktů – morfogenů (Vesmír 74, 76, 1995/2). Při vzniku a organizaci morfogenetických polí mají významnou úlohu právě homeoboxové geny. Tak v základech končetin vznikají gradienty proteinů Hox, které v určitých místech indukují produkci specifických proteinů a tak vytvářejí podmínky pro vznik pole včetně jeho polárních os (obr. obrázek). 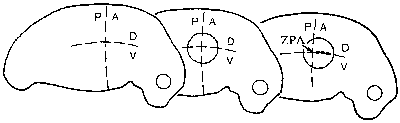 Končetiny obratlovců a hmyzu mají zcela odlišnou stavbu a byly vždy považovány za analogické struktury, tj. struktury různého původu, ale přizpůsobené témuž účelu. Molekulárněgenetická analýza ukázala, že stejné proteiny, které se uplatňují při vzniku nohy hmyzu z imaginálního terčku 1) , představujícího morfogenetické pole, se podílejí i na vzniku končetiny obratlovců.
Končetiny obratlovců a hmyzu mají zcela odlišnou stavbu a byly vždy považovány za analogické struktury, tj. struktury různého původu, ale přizpůsobené témuž účelu. Molekulárněgenetická analýza ukázala, že stejné proteiny, které se uplatňují při vzniku nohy hmyzu z imaginálního terčku 1) , představujícího morfogenetické pole, se podílejí i na vzniku končetiny obratlovců.
{kind=link}
V různých morfogenetických polích působí shodné produkty řídících genů na různé, specifické cílové geny. Primárním polem je celé embryo v průběhu blastogeneze, časného vývoje, které se postupně rozpadá na různá morfogenetická pole orgánových základů. Morfogenetická pole jsou založena na geneticky definovaných interakcích mezi buňkami a hranice jejich kompetence může být dána homeotickými geny. Evoluce závisí na replikaci a modifikaci morfogenetických polí. Morfogenetické pole sjednocuje atomizmus genetických a biochemických drah v celkovost drah vývojových. Tak jako buňka (a nikoli přímo geny) je jednotkou struktury a funkce těla, tak je morfogenetické pole základní jednotkou ontogenetických a fylogenetických změn. To ovšem neznamená, že geny snad nejsou pro vývoj důležité, ale jejich produkty musí vzájemně reagovat. Pole představují modulární povahu vyvíjejícího se embrya, zprostředkují vztah mezi genotypem a fenotypem.
Literatura
De Robertis E. M., Sakai Yoshiki: A common plan for dorsoventral patterning in Bilateralia. Nature 380, 37 – 40, 1996.Gilbert S. F., Opitz J. M., Raff R. A.: Resynthesizing evolutionary and developmental biology. Develop. Biol. 173, 357 – 372, 1996.
Valentine J. W., Erwin D. H., Jablonski D.: Developmental evolution of Metazoan bodyplan: the fossil evidence. Devel. Biol. 173, 373 – 381, 1996.
Poznámky
V posledních letech se toulá po světě spasitel. Nenápadnými kradmými kroky se už zase blíží milénium, není tedy nijak zvláště podivné, že se po světě toulají spasitelé. Ten přírodozpytcův je nazván svými učedníky molekulární biologie. Každým dnem prostřednictvím svých nadpřirozených metodických postupů páchá zázraky na denním světle a bez uzardění slibuje odpovědět na (takřka) vše, co jen neukojeného človíčka napadne. Tento mesiáš je skvělý, je vášnivý, je neopakovatelný, je nejlepší.
Je ovšem zábavné na každém neposkvrněném početí hledat cosi nepatřičného, cosi zasmrádajícího, naše punkerská generace je zkrátka taková. Už nějaký čas přinejmenším pár lidí po světě nevěří na to, že by fylogenetické analýzy i těch nejkonzervativnějších sekvencí ribozomálních nukleových kyselin (tím spíš takových těch o něco míň konzervativních sekvencí, jako je rbcL nebo ubiquitin) samy o sobě rozlouskly záhady všeho stvoření a definitivně postulovaly ty pravé, niterné vztahy mezi organizmy. Naopak, stále sílí tendence pokoušet se při fylogenetických analýzách o totální evidenci, tedy bezbožné použití molekulárních i morfologických dat v jedné veliké, všepohlcující matici znaků, která se následně rozžvýká příslušným metodickým postupem.
Jiným velkým zázrakem je používání analýz sekvencí homeoboxových genů při hledání svatého grálu evolučních a systematických biologů, totiž – homologií. Všem je jasné, proč je nutné homologie hledat, zhusta to i sem tam někdo činí, ale ony se, mrchy, bohužel hledají dost špatně. Homeoboxové geny jsou ale přece zázrak nade všechny ostatní! Je tak elegantní vyvodit homologii mezi komplexy morfologických znaků (výhradně) na základě příbuznosti homeoboxových genů hrajících klíčové role v jejich utváření během ontogeneze.
Pýcha často předchází pád. Buďme tedy obezřetní a společně s panem Frietsonem Galisem (viz říjnové číslo časopisu Trends in Ecology and Evolution – TREE) se podívejme na celou věc blíž. Nemělo by naše úvahy zneklidňovat vědomí stovek milionů let samostatné evoluce taxonů, jejichž struktury chceme homologizovat? Přes nepochybně velkou konzervativnost homeoboxových genů existuje určitá možnost, že mohlo dojít k nějakým tvrdším změnám ve vztahu mezi geny a příslušnými morfologickými strukturami. Objevují se názory, že je přinejmenším nerozumné homologizovat různé struktury na základě molekulárních dat bez pokusu o vhled do jejich vývojových mechanizmů.
Především by mělo být důležité nějak doložit, že homologní homeoboxové geny jsou účastny ve formování skutečně homologních struktur. Při srovnávání různých skupin organizmů se zdá, že je snazší nalézt příklady homologních homeoboxových genů spojených spíše s nehomologními než homologními strukturami. Ukázkou budiž obratlovčí gen Brachyury (T) zapojený ve vývoji hřbetní struny a jeho hmyzí obdoba gen T angažovaný ve formování zadního střeva. Podobná situace se opakuje i po sestupu poněkud níže v hierarchii tvorstva – například v rámci hmyzu. Gen Distalless se v drozofile podílí na předozadní organizaci končetin a křídel dospělců, naproti tomu u motýla Precis coenia je tento gen exprimován v panožkách housenek a u dospělců v oblasti křídel v místech, kde se zakládají taková ta pěkná oka. Panožky by měly být homologní s normálními končetinami – potud je všechno v pořádku. Fleky na křídlech v podobě ok by měly být homologní s nohama nebo křídly už o dost míň. Příklady, kdy jsou homologní geny zřetelně zapojeny ve formování homologních struktur a ne toliko v určení podobné poziční informace, se nacházejí pouze uvnitř hmyzu nebo uvnitř obratlovců.
Proč tedy obvykle nejsou homologní geny účastny v zakládání homologních struktur? Pan Galis nabízí několik různých vysvětlení:
- Lze předpokládat, že duplikace homeoboxových genů (či jejich skupin) zřejmě vedla ke vzniku nadbytečné genetické informace. Některé práce ukazují, že tato redundantní informace mohla být v nezávisle se vyvíjejících liniích nezávisle použita, což mohlo vyústit v zapojení homologních genů do vývoje nehomologních struktur.
- Stejné homeoboxové geny mohou signalizovat stejnou poziční informaci, a mít tedy stejnou funkci, ale jimi oslovené buňky mohou založit nehomologické struktury. Evoluční změna se v takovém případě objevuje někde na hierarchicky nižší hladině, než je homeoboxový ovládač.
- Pleiotropní (tedy mnohofunkční) homeoboxové geny mohly během evoluce ztrácet své jednotlivé funkce, přičemž tyto ztráty mohou být samozřejmě u různých linií potomstva různé.
- Ontogeneze je interaktivní proces. Funkce homeoboxových genů závisí na kontextu, ve kterém jsou exprimovány. Proto by měl být při hledání homologií nutně brán zřetel na vývojové mechanizmy. Evoluční změny mění vývojové mechanizmy, novinky ve struktuře se uskutečňují skrze změny ve vývojových interakcích.
Vezmeme-li například obratlovce a hmyz, jejich ontogeneze se v některých ohledech dosti liší, zejména pokud jde o chování tkání. U členovců jsou tkáně tvořeny plochami vzájemně spojených buněk, tedy epitely, které se vyvíjejí v podstatě autonomně. Vývoj obratlovců naproti tomu kromě vývoje epitelů zahrnuje i spolupráci mezenchymu, jehož izolované buňky migrují do lokálních epitelů, s nimiž následně interagují. Ontogeneze u obratlovců navíc probíhá prostřednictvím kaskády induktivních procesů. Zdá se tedy značně obtížné hledat homologie zrakových orgánů hmyzu a obratlovců za situace, kdy k jejich vzniku dochází zcela odlišnou cestou.
Genetické mutace se mohou přihodit na každé úrovni hierarchické organizace vývoje organizmu a evoluční novinky způsobené jejich účinkem se mohou objevit na kterékoli z těchto úrovní. Nepatrné mutace mohou mít nepatrný vliv, anebo taky vliv ohromný, plynoucí z celé kaskády interakcí. To je podle Galise příčina toho, proč je tak problematické hledat pomocí homeoboxových genů homologie struktur například u obratlovců a u hmyzu. A co hůř – proč je celý tento koncept homologií tak obtížně interpretovatelný a v posledku tak kontroverzní.
Tak především, zamyslete se nad tím, co jest homologie (podrobněji viz článek J. Zrzavého, Vesmír 76, 27, 1997/1). Dále, nevěšte se. Stejně se neustále motáme na různých hierarchických hladinách organizace živých soustav a tak nějak nevíme, co je to znak, a přitom právě v tom nám mohou ontogenetické mechanizmy leccos napovědět. A v posledku považte: to, že jsme humři vzhůru nohama, je přece neuvěřitelně nádherné.
Stanislav Mihulka
Galis F.: The evolution of insects and vertebrates: homeobox genes and homology. TREE 11:10, 402–403, 1996
O autorovi
František Sládeček
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
