










Hromadná vymírání bořící a tvořící
| 5. 8. 1994 Geologická historie života byla poznamenána řadou krizí. Ty, které vynikají mimořádnou ničivostí a rychlostí, označujeme jako katastrofy. Biologické katastrofy se vyznačují celosvětovým pronikavým snižováním diverzity a vymíráním dosud prosperujících vývojových linií. Ekologický prostor uvolněný vymíráním obsazují skupiny organizmů, které dosud byly v pozadí. Mění se taxonomické složení jednotlivých úrovní potravních řetězců, mění se i vývojové trendy. Smrt je pro existenci druhů stejně důležitá jako zrození. V proměnlivých životních podmínkách umožňuje růst nové, fenotypově adaptované generace, a tím zvýšení pravděpodobnosti dalšího přežití druhu. V historii vývojových linií je do určité míry obdobou smrti jedinců vymírání druhů nebo vyšších taxonů. Proto považují někteří evolucionisté mezidruhovou selekci za základní makroevoluční mechanizmus, který eliminací jedněch druhů a diverzifikací jiných vede během geologického času ke vzniku vývojových trendů. Vymírání tedy na jedné straně „boří“ a na druhé se spoluúčastní formování cest evoluce života. Během nesčetných epizod vymírání, které lze předpokládat v rané historii vývoje života, se například prosadilo jako výhodná strategie přežívání mnohobuněčných organizmů pohlavní rozmnožování. Přesto, že ve srovnání s rozmnožováním nepohlavním znamená pro organizmy značnou časovou i energetickou zátěž, přináší pohlavní rozmnožování, kromě jiných výhod, i schopnost lépe uchovávat velkou mnohotvárnost (polymorfizmus) jedinců v rámci druhu. A to je právě jeden z důležitých faktorů zvyšujících jak šanci druhů na přežití během změn prostředí vedoucích k vymírání, tak pravděpodobnost obsazení uvolněného ekologického prostoru po vymírání.
Geologická historie života byla poznamenána řadou krizí. Ty, které vynikají mimořádnou ničivostí a rychlostí, označujeme jako katastrofy. Biologické katastrofy se vyznačují celosvětovým pronikavým snižováním diverzity a vymíráním dosud prosperujících vývojových linií. Ekologický prostor uvolněný vymíráním obsazují skupiny organizmů, které dosud byly v pozadí. Mění se taxonomické složení jednotlivých úrovní potravních řetězců, mění se i vývojové trendy. Smrt je pro existenci druhů stejně důležitá jako zrození. V proměnlivých životních podmínkách umožňuje růst nové, fenotypově adaptované generace, a tím zvýšení pravděpodobnosti dalšího přežití druhu. V historii vývojových linií je do určité míry obdobou smrti jedinců vymírání druhů nebo vyšších taxonů. Proto považují někteří evolucionisté mezidruhovou selekci za základní makroevoluční mechanizmus, který eliminací jedněch druhů a diverzifikací jiných vede během geologického času ke vzniku vývojových trendů. Vymírání tedy na jedné straně „boří“ a na druhé se spoluúčastní formování cest evoluce života. Během nesčetných epizod vymírání, které lze předpokládat v rané historii vývoje života, se například prosadilo jako výhodná strategie přežívání mnohobuněčných organizmů pohlavní rozmnožování. Přesto, že ve srovnání s rozmnožováním nepohlavním znamená pro organizmy značnou časovou i energetickou zátěž, přináší pohlavní rozmnožování, kromě jiných výhod, i schopnost lépe uchovávat velkou mnohotvárnost (polymorfizmus) jedinců v rámci druhu. A to je právě jeden z důležitých faktorů zvyšujících jak šanci druhů na přežití během změn prostředí vedoucích k vymírání, tak pravděpodobnost obsazení uvolněného ekologického prostoru po vymírání.
Vymření může být fyletické, během něj se jeden taxon transformuje v jeden nebo více jiných taxonů (pseudoextinkce – taxon vlastně nevyhyne, i když vymizí), nebo vymření bez potomků, typické právě pro období krizí. V geologické historii se střídají delší období s malou frekvencí vymírání s relativně krátkými obdobími zvýšené intenzity vymírání, do kterých jsou v geologické časomíře často kladeny hranice geochronologických jednotek (věků, epoch, ér). Jednotlivá vymírání se od sebe liší množstvím postižených druhů, rozmanitostí jejich ekologických tolerancí i jejich postavením v ekosystémech. Od běžných vymírání se mimořádnou intenzitou liší hromadná vymírání. Nejčastěji se definují jako relativně krátkodobá událost (1 – 4 miliony let), během které vymře více než 50 % druhů v několika geograficky široce rozšířených diverzifikovaných vývojových liniích (vyšších taxonech). Žádné masové vymírání neproběhlo v jediném krátkém okamžiku, jak bychom mohli intuitivně předpokládat z pojmu katastrofa.
Zvýšený zájem o studium příčin a důsledků krizí v biosféře je v posledních zhruba 15 letech živen jednak rychlou devastací životního prostředí, v mnoha důsledcích vyvolávající stejné efekty jako hromadná vymírání v geologické historii, jednak atraktivní hypotézou o srážce naší planety s kosmickým tělesem, která měla vést na hranici křídy a třetihor k vymření mnoha skupin organizmů, včetně dnešních miláčků dětí – dinosaurů.
Příčiny vymírání
Možných příčin vymírání je ještě více než možných příčin smrti. Přímo na organizmy působí bezprostřední příčiny, namátkou uveďme rychlou změnu teploty nebo salinity, zánik vhodného biotopu apod. Ty mají své vlastní příčiny, označované jako prvotní. Jedna bezprostřední příčina může být vyvolána různými prvotními příčinami. Globální ochlazení může být důsledkem jak změny oceánické cirkulace vyvolané změnou postavení kontinentů, tak nástupem intenzivní vulkanické činnosti. Příčiny vymírání mohou být jednak biotické (konkurence, nemoci, vyhubení masožravci), jednak abiotické. Biotickými příčinami lze vysvětlit snad jen některá menší vymírání. Změny prostředí, konkrétně ochlazení ve čtvrtohorách a rozšíření stepních formací, hrály významnější roli než konkurence i při vymizení většiny jihoamerických vačnatců po imigraci placentálních savců ze Severní Ameriky před 3 miliony let. Hromadná vymírání vyvolávaly spíše události abiotické, na které biotické vlivy navazovaly jako druhotné. Do výčtu nejčastěji uvažovaných abiotických příčin hromadných vymírání patří:- klimatické změny,
- redukce rozlohy mělkých moří na kontinentálních blocích a jejich okrajích v době globálního (eustatického) poklesu hladiny oceánů,
- lokální změny obsahu solí v mořské vodě,
- mocná vrstva anoxické (na kyslík chudé) vody v oceánech zaplavující šelfy a epikontinentální moře, kde dusí organizmy žijící při dně,
- pokles provincionality (splynutí kontinentů, vymizení bariér),
- zvýšená sopečná činnost (popel v atmosféře a aerosoly vzniklé při vysokém obsahu SO2 snižují průchodnost atmosféry pro sluneční záření a dochází k ochlazení, deště rozpouštějící SO2 a CO2 okyselují svrchní partie moří, HCl v atmosféře může redukovat ozonovou vrstvu, převaha CO2 ve vulkanických plynech může vyvolat oteplení mechanizmem skleníkového efektu),
- zvraty polarity magnetického pole Země, během kterých by oslabený magnetický štít umožnil dopad kosmického záření na zemský povrch,
- dopad mimozemského tělesa nebo jiné mimozemské vlivy (výbuch supernovy, průchod sluneční soustavy mračnem kosmického prachu atd.).
Bouřlivou diskusi vyvolal předpoklad periodicity vymírání, který před deseti lety zveřejnili David Raup a John Sepkoski z Chicagské univerzity. Pravidelné prudké zvyšování počtu vymření vždy po 26 milionech let, počínaje koncem prvohor, by mělo mít podle nich nejspíše kosmické příčiny, např. periodické bombardování kometami vychylovanými z Oortova oblaku vlivem pravidelného přibližování hypotetického průvodce Slunce, malé hvězdy Nemesis, nebo oscilaci sluneční soustavy kolem galaktického rovníku s větší koncentrací hmoty. Po podrobnější analýze užitých dat a metod se ukazuje, že o striktní pravidelnosti nelze uvažovat. Určitá kvaziperiodicita by však mohla podle některých názorů navazovat na procesy probíhající uvnitř Země. Motorem by mělo být teplo stoupající ze zemského jádra a zachycující se na bázi zemského pláště. Postupně narůstající mocnost takto vznikající silně prohřáté vrstvy by měla vést ke vzniku termických sloupců, míst, kde z hlubin pláště stoupá k povrchu horké magma. Je to jako když se vaří v hrnci voda a v nejteplejších místech stoupá vzhůru. Po dlouhé době dorazí magma až k povrchu a rozlévá se za vzniku ohromných čedičových příkrovů. Výlevy magmatu fungují jako ventil – po patřičném zmenšení mocnosti přehřáté vrstvy na bázi pláště termické sloupce přestávají fungovat a začíná nový cyklus.
Hromadná vymírání
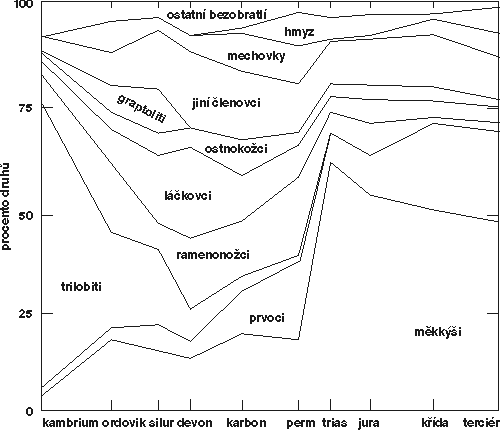
Každé hromadné vymírání bylo jedinečné – jiné oběti, jiná kombinace příčin, jiný průběh. Od počátku prvohor, kdy nastal rozvoj organizmů s pevnými schránkami vhodnými pro fosilizaci, proběhlo pět epizod hromadného vymírání. První je krize na hranici ordoviku a siluru (O/S) před 438 miliony let. Vymřelo 22 % čeledí mořských bezobratlých. Nejvíce byly postiženi trilobiti, graptoliti, ramenonožci, koráli a mechovky, značně byly eliminovány útesová společenstva a plankton. Za všechno zřejmě mohlo zalednění Gondwany, jižního superkontinentu ležícího tehdy v okolí jižního pólu. Počátek glaciace vyvolal první fázi vymírání, která nastala 1 – 2 miliony let před koncem ordoviku. Zadržení velké kubatury vody v kontinentálních ledovcích způsobilo snížení hladiny oceánů. Na hranici O/S naopak ledovce rychle tály. Začal rychlý celosvětový vzestup hladiny oceánů a v mnoha oblastech nastoupily anoxické podmínky (nedostatek kyslíku). Další hromadné vymírání postihlo především organizmy útesových ekosystémů a proběhlo před 367 miliony let, v mladším devonu. Kromě většiny útesotvorných korálů vymřela především řada skupin mořského fytoplanktonu (až 90 % druhů), zooplanktonu, ramenonožců, amonitů, pancířnatých a trnoploutvých ryb. Sladkovodní ryby byly naproti tomu postiženy jen málo. Různé skupiny vymíraly po etapách po dobu 3 – 4 milionů let. Příčiny nejsou dosud příliš jasné. Několikrát se objevily údaje o zvýšeném obsahu iridia poblíž této hranice. Po ruce byla impaktní hypotéza – předpoklad dopadu velkého asteroidu. Její slabinou však je, že iridiová anomálie vznikla až asi 1,5 milionu let po hromadném vymírání. Větší pozornost tedy zaslouží možné pozemské příčiny. Postižení převážně teplomilných skupin ukazuje, že příčinou vymírání mohlo být ochlazení. V geologickém záznamu se skutečně objevuje obdoba událostí z hranice O/S: pokles hladiny následovaný po krátké době jejím vzestupem a rozšířením anoxického režimu. Předpoklad ochlazení potvrzují stopy svrchnodevonského zalednění zjištěné v Jižní Americe.
Další hromadné vymírání postihlo především organizmy útesových ekosystémů a proběhlo před 367 miliony let, v mladším devonu. Kromě většiny útesotvorných korálů vymřela především řada skupin mořského fytoplanktonu (až 90 % druhů), zooplanktonu, ramenonožců, amonitů, pancířnatých a trnoploutvých ryb. Sladkovodní ryby byly naproti tomu postiženy jen málo. Různé skupiny vymíraly po etapách po dobu 3 – 4 milionů let. Příčiny nejsou dosud příliš jasné. Několikrát se objevily údaje o zvýšeném obsahu iridia poblíž této hranice. Po ruce byla impaktní hypotéza – předpoklad dopadu velkého asteroidu. Její slabinou však je, že iridiová anomálie vznikla až asi 1,5 milionu let po hromadném vymírání. Větší pozornost tedy zaslouží možné pozemské příčiny. Postižení převážně teplomilných skupin ukazuje, že příčinou vymírání mohlo být ochlazení. V geologickém záznamu se skutečně objevuje obdoba událostí z hranice O/S: pokles hladiny následovaný po krátké době jejím vzestupem a rozšířením anoxického režimu. Předpoklad ochlazení potvrzují stopy svrchnodevonského zalednění zjištěné v Jižní Americe.
Nejvíce byla biota v pokambrickém období zdecimována před 250 miliony let na hranici permu a triasu (P/T). V mořích vymřelo 80 – 95 % druhů s pevnými schránkami a kostrami. Vymírání probíhalo s proměnlivou intenzitou v intervalu maximálně 5 milionů let, hlavní pulz pak pouze jeden milion či méně. Zanikla prvohorní společenstva přisedlých filtrátorů mělkého mořského dna, hlavně ramenonožců, lilijic, poupěnců, mechovek a prvohorních typů korálů. Vymřelo mnoho zooplanktonu i velkých predátorů – prvohorních skupin amonitů a ryb. Definitivně zmizely i poslední zbytky trilobitů. Na souši podlehlo krizi 8 z 27 tehdejších řádů hmyzu, jejichž existenci se podařilo doložit. Naproti tomu mezi obojživelníky a sladkovodními rybami nebyly zaznamenány žádné významné změny. Vymírání plazů má maximálně charakter průběžného vymírání, stejně jako postupná výměna plavuňovitých, přesličkovitých, kapradinovitých a kapraďosemenných rostlin za jehličnany vyvolaná nástupem suššího klimatu v tropických oblastech obřího superkontinentu Pangey. Při vysvětlování příčin této krize se musí počítat s kombinací různých událostí, říká odborník na tuto hranici Douglas Erwin. Prvotní příčinou krize by mohlo být zalednění v důsledku intenzivní vulkanické činnosti, která byla spojena se vznikem sibiřských trapů, rozsáhlých plochých výlevů čedičů mezi Lenou a Jenisejem o kubatuře zhruba 2 miliony km3. Radiometrické údaje o jejich stáří se velmi dobře kryjí s dobou tohoto masakru. V geologickém záznamu se opět objevují indicie poklesu hladiny oceánu (o 150 až 210 m), vzhledem k rychlosti nejspíše vyvolaného nahromaděním vody v ledovcích, a následně rychlého vzestupu provázeného rozšířením anoxických podmínek. Bezprostředními příčinami vymírání by mohly být zpočátku hlavně ochlazení a zvětšení sezónních teplotních rozdílů. Hlavní zásah do mořských ekosystémů by pak mohl spadat do konce doby ledové. Jestli totiž konfigurace kontinentálních bloků podporuje atmosférickou a oceánickou cirkulaci určující teplé klima, pak skončení epizody extrémního vulkanizmu povede k rychlému oteplení a rychlému tání ledovců. Tím se uvolní do oceánů velké množství sladké vody. Snížení salinity svrchních částí oceánů postihne obvykle plankton, který je na tyto změny citlivý, planktonické larvy a aktivně plavající větší živočichy. Protože voda s menší salinitou je méně hustá, vytváří u hladiny „pokličku“ bránící klesání prokysličené vody do hloubky. Anoxické podmínky pak způsobí udušení všech na dně žijících organizmů kromě anaerobních bakterií. Je zajímavé, že tento katastrofický scénář lze užít k vysvětlení základních rysů všech hromadných vymírání. Dosud však nejsou pro každé z nich doklady o existenci stejně starého rozsáhlého zalednění.
 Čtvrté masové vymírání v pokambrické době zasáhlo biotu koncem triasu. Z bohatě diverzifikovaných triasových amonitů vymřelo 58 čeledí, postiženi byli mlži, břichonožci, ramenonožci a mořští plazi. Na souši vyhynulo 35 čeledí hmyzu a velká změna nastala mezi plazy. Některé skupiny zmizely úplně, jiné byly značně zredukovány. V tomto případě je pro fanoušky mimozemských příčin v Kanadě k dispozici meteorický kráter Manicouagan o průměru 70 km, vzniklý v intervalu mezi 206 a 213 miliony let, kam spadá i hranice triasu a jury (210 milionů let). V geologických profilech však nebyly v této úrovni nalezeny ani mikrotektity, ani iridiová anomálie. Na hranici triasu a jury vymíraly hlavně mořské organizmy. Suchozemští plazi byli intenzivně postiženi již o 12 – 17 milionů let dříve. Jejich vymírání souviselo s klimatickými změnami (aridizací) a náhlou změnou rostlinstva.
Čtvrté masové vymírání v pokambrické době zasáhlo biotu koncem triasu. Z bohatě diverzifikovaných triasových amonitů vymřelo 58 čeledí, postiženi byli mlži, břichonožci, ramenonožci a mořští plazi. Na souši vyhynulo 35 čeledí hmyzu a velká změna nastala mezi plazy. Některé skupiny zmizely úplně, jiné byly značně zredukovány. V tomto případě je pro fanoušky mimozemských příčin v Kanadě k dispozici meteorický kráter Manicouagan o průměru 70 km, vzniklý v intervalu mezi 206 a 213 miliony let, kam spadá i hranice triasu a jury (210 milionů let). V geologických profilech však nebyly v této úrovni nalezeny ani mikrotektity, ani iridiová anomálie. Na hranici triasu a jury vymíraly hlavně mořské organizmy. Suchozemští plazi byli intenzivně postiženi již o 12 – 17 milionů let dříve. Jejich vymírání souviselo s klimatickými změnami (aridizací) a náhlou změnou rostlinstva.
Dosud poslední, nejmladší hromadné vymírání bylo před 65 miliony let na hranici křídy a třetihor (K/T). Jistě ale posledním nezůstane. Je možné, že budoucí badatelé (budou to lidé ?) vyhodnotí současně probíhající vymírání jako hromadné. Vzhledem k současnému rozsahu a tempu narušování důležitých složek globálního ekosystému lidskou společností je to dokonce dost pravděpodobné (viz také tabulku III ve Vesmíru 72, 524, 1993/9).
Protože diskuse o příčinách K/T krize, při které vymřelo 60 – 75 % druhů, je dosud velmi živá, shrňme pouze současné skóre zápasu mezi zastánci dopadu asteroidu a obhájců názoru, že na to, co se stalo, stačily i pozemské síly. Ve prospěch impaktní teorie se uvádí:
- deseti až stonásobné obohacení hraniční vrstvičky iridiem, kovem vzácným v zemské kůře, zato běžným v asteroidech,
- nálezy mikrotektitů, drobných kuliček vzniklých roztavením hornin v místě dopadu a rozstříknutých do ohromných vzdáleností,
- výskyt šokových mikrostruktur křemene, které mohou vzniknout jen při extrémních tlacích, jaké nastávají právě při dopadu velkého kosmického tělesa,
- výskyt chaotických usazenin vzniklých působením vln tsunami, vysokých stovky metrů,
- vysoký obsah sazí svědčící o ohromných požárech,
- téměř okamžité vymření vápnitého planktonu. Posledním trumfem impaktistů byl nález kráteru Chicxulub na šelfu severně od poloostrova Yukatan, vzniklého údajně v době krize. Přestože má mít v průměru asi 150 – 180 km, což odpovídá předpokládanému průměru asteroidu okolo 10 km, morfologicky se neuplatňuje, protože se nachází v moři a je zakryt mladšími sedimenty. Odpůrci impaktu se snaží vysvětlit každý argument impaktistů pozemskými příčinami – iridium vynášené z hlubokých částí zemského pláště vulkanickou činností, mikrotektity jako vulkanické částice nebo špatně interpretované minerální výplně kulovitých schránek jednobuněčných planktonických řas vzniklé během zpevňování sedimentů, vznik šokových křemenů extrémně silnými sopečnými výbuchy atp. Jejich silnou zbraní je paleontologická evidence. Většina skupin, které vymřely – dinosauři, amoniti, tlustoskořepatí mlži rudisti – začala mizet v několika etapách již dlouho před K/T hranicí. Stupňovité vymírání, napřed nejteplomilnějších druhů, a teprve pak i druhů „otužilejších“, ukazuje na postupné ochlazování. Ponenáhlý pokles hladiny oceánů koncem křídy o 80 – 130 m může svědčit o ochlazování doprovázeném tvorbou kontinentálních ledovců; pro ty ale chybějí přímé doklady. Množí se pozorování mikropaleontologů zjišťující, že ani rychlá decimace planktonických mikroorganizmů neproběhla ze dne na den, ale trvala několik set tisíc let. Začala již před usazením vrstvičky s iridiovou anomálií a přednostně vymíraly skupiny citlivější ke změnám salinity. Podobnost se strukturou vymírání na konci permu je značná. Globální ochlazení se dá přitom velmi dobře vysvětlit jako důsledek ohromné vulkanické činnosti. Právě v době K/T krize se v západní Indii vylilo během zhruba milionu let minimálně 1,5 milionu km3 čedičů, které vytvořily Dekanskou plošinu o rozloze zhruba 100 000 km2 s celkovou mocností vulkanických produktů 1 200 až 1 800 m. Silné argumenty tedy nechybějí ani jednomu táboru. Která názorová skupina má pravdu, je dosud těžké rozhodnout. Určitou možnost nabízí řešení kompromisní – asteroid dopadl v době, kdy právě probíhalo jedno z větších vymírání vyvolaných klimatickou změnou, a “dorazil“ to, co ochlazování začalo. V poslední době se mi však zdá, že pozorovanému průběhu vymírání lépe odpovídá teorie vulkanická. Shrnutí současných názorů na příčiny hromadných vymírání tedy dává za pravdu již vousaté hypotéze, že to byly hlavně změny klimatu, ať již byly vyvolány čímkoli. Ekologickou rozmanitost jejich obětí pak měla na svědomí řada událostí, včetně biotických, které na tyto klimatické výkyvy kaskádovitě navazovaly.
Proč vymřeli dinosauři?
Vymření takových bohatých skupin, jako byli dinosauři nebo trilobiti, vyvolává řadu otázek. Vytlačila je jiná, lépe přizpůsobená skupina? Vymřeli náhodou v důsledku kosmické katastrofy? Nebo měli „špatné geny“? Odpověď není jednoduchá a ty, kteří doufají, že se někdy najde jednoznačné řešení, asi neuspokojí.Vymírání diverzifikovaných vyšších taxonů je vždy složitý historický proces, který je výsledkem dlouhodobého nepoměru mezi vymíráním bez potomků a odštěpováním nových druhů. Když převažuje odštěpování nových druhů, skupina se rozrůzňuje a počet druhů stoupá, v opačném případě diverzita klesá a vývojová linie po určité době vymře. Důležité je, že druhy nemizí pouze během hromadných vymírání. Naopak, např. 96 % mořských bezobratlých ukončilo svou existenci při běžných vymíráních.
Ani všechny vývojové linie řazené do skupiny Dinosauria nevymizely najednou na konci druhohor. Některé byly uťaty již v juře, např. stegosauři, jiné, a těch byla většina, ještě před koncem křídy. Samotné hranice křídy a třetihor se podle paleontologických záznamů dožil sotva tucet rodů. Lze se domýšlet, že například pro obří druhy dinosaurů byly během krizí problémem jejich velké rozměry. Nemohli se nikam schovat a potřebovali příliš mnoho potravy. Významná byla i malá populační hustota, menší počet druhů v populacích a dlouhá doba obměny generací. Výhodnost větších rozměrů pro jedince se uplatňovala jako selekční faktor v běžných dobách, v době krize převážila nevýhodnost extrémní velikosti pro přežití druhu. Pro velké i malé druhy mohla hrát důležitou roli citlivost k snížení teploty v době inkubace vajec, až na výjimky asi nedostatečně kompenzovaná „tepelnou“ péčí o snůšku (ptáci na vejcích sedí a jako jediní z dinosaurovitých dodnes přežili – viz následující článek Zbyňka Ročka). Pro býložravé druhy byla významná změna typu vegetačního krytu, pro velké dravé druhy ubývání velkých býložravců. A to jistě nejsou zdaleka všechny možnosti zúčastněných příčin. Připomeneme-li si složitý propletenec změn prostředí probíhajících během K/T krize, je zřejmé, že univerzální vysvětlení vymření dinosaurů jednou bezprostřední příčinou neexistuje. Každý druh dinosaurů byl adaptován na trochu odlišné podmínky a vyhynul pravděpodobně z poněkud jiných příčin.
Stejně jako vymírání nepřizpůsobených druhů v době rychlé změny prostředí může k vymizení vyšších taxonů vést i ztráta evoluční plasticity vývojové linie. Označuje se tak soubor vlastností umožňujících rychle vytvářet nové druhy nebo pronikat do nových adaptivních zón. Přispívají k ní modifikovatelnost morfologických struktur k novým účelům (preadaptace), fyziologická flexibilita umožněná např. zdvojením některých genů, mutabilita, schopnost porušit koordinovanost růstu jednotlivých částí těla během ontogeneze (heterochronie) apod. Evoluční plasticita umožňuje vývojovým liniím, které přežily nějakou krizi, rychle „vysílat výhonky“ do uvolněného ekologického prostoru. Jako častá příčina snížení evoluční plasticity bývá uváděna přílišná specializace na určitý typ životního prostředí, potravní specializace nebo vznik extrémních tvarů podmíněný pohlavním výběrem. Jeden vyšší taxon může postupně vymřít a být v ekosystému nahrazen jiným i v případě, že v průběhu více krizí dopadnou oba taxony stejně špatně, tj. v obou vymře stejný počet druhů. Rozhodující pak není odolnost jednotlivých vývojových linií v době krizí, ale schopnost rychle se chopit příležitosti, rychle obsadit uvolněný ekologický prostor. Zatímco jeden taxon se v takovém případě postupně redukuje, druhý se rozrůstá, diverzifikuje a zvyšuje tak pravděpodobnost svého přežití v další krizi. Je zajímavé, že rychlost tohoto typu evoluce je určována rychlostí vymírání.
Obecnou otázku „Proč vymřeli dinosauři?“ lze tedy zodpovědět také jen obecně: Parafyletický taxon dinosauři (do kterého neřadíme z dinosaurů vzniklé ptáky) vymřel proto, že změny prostředí byly moc rychlé, různorodé a intenzivní a mezi dinosaury nebyl žádný druh, který by vydržel změny životních podmínek, ani druh, který by se těmto změnám dokázal dostatečně rychle přizpůsobit. To, že takové druhy mezi dinosaury koncem křídy chyběly, lze vysvětlit dlouhodobým hromaděním různých specializací vedoucích k postupnému snižování jejich evoluční plasticity.
Literatura
Vladimír Pokorný a kol.: Všeobecná paleontologie, UK, Praha 1992Obrázky








{kind=link}
{kind=link}
{kind=link}
Citát
Miroslav Holub, O příčinách porušení a zkázy těl lidských, Pražská imaginace, Praha 1992, str. 135
Jatky jsou jakýmsi nočním a spodním vědomím města, jež na jedné straně potřebuje a žádá živočišné bílkoviny za příčinou zdravého vývoje mladých i starých generací, na druhé straně však si ekluje páchnoucí krev, střeva s mezentériem a reflexní záškuby svalů. Pro slušného člověka jsou smrtelné křeče, cožkoli se tak nazývá, přijatelny u much, myší, kuřat a kaprů, nepřijatelny však u velkých savců s temnýma moudrýma, nebo docela smutnýma očima. Krev je přijatelná ve stíratelných kapkách na kuchyňském stole, nikoli v loužích a brodech na betonové podlaze.
‘Já bych to dělat nemoh,’ praví slušný člověk.
Slušný člověk ostatně omdlévá i v pitevně, vyspělejší individua i v sále operačním.
Morální kodex delimitovaný výrazem já-bych-to-dělat-nemoh a omdléváním jest ovšem morálním kodexem tartuffovským. Když slyším vysoce kulturní konstatování já-bych-to-dělat-nemoh, říkám si v duchu ale moh, ale moh, kdyby se za to vzalo; vzít za to, to jest vzít za člověka, umí každý pořádný policajt, zejména je-li mezi dalšími normálními policajty, jednak to ukazuje známý psychologický pokus s dobrovolníky, kteří se kultura – nekultura a kodex – nekodex dají v experimentální situaci donutit k působení krutých bolestí člověku za stěnou...
O autorovi
Jan Krhovský
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
