









Lužné lesy a osud ich fauny
Lužné lesy predstavujú súbor ekosystémov, výskyt a zloženie ktorých určuje hladina spodnej vody. Vyskytujú sa tam, kde spodná voda zasahuje do koreňovej vrstvy stromov alebo sa k nej aspoň silne približuje a kde dochádza aspoň k občasným záplavám. Spravidla na jar, ale pri Dunaji aj viackrát do roka. Lužné lesy sa preto rozprestierajú v plochom teréne v blízkosti vodných tokov a ich ramien. Podľa hladiny spodnej vody a pravideľnosti a dĺžky trvania záplav sa fytocenologicky delia na dve skupiny.
Prvú tvoria porasty zložené z drevín znášajúcich dlhodobé zaplavenie a podmáčanie. Teda hlavne z rôznych druhov vŕb, topoľov a jelší. Zlatníkova škola ich zaraďuje do skupiny geobiocénov vŕbové jelšiny (Salici alneta), Montpelierská škola do zväzu Salicion. Vyskytujú sa bezprostredne pri vodných tokoch a plochách, kde ich zastupuje hlavne asosiácia Salici-Populetum (vŕbovo-topoľové lesy). Konvenčne sa označujú ako tzv. mäkké luhy.
Druhú skupinu tvoria porasty na periodicky krátkodobo zaplavovaných miestach, kde spodná voda zasahuje len po určitú časť roka do koreňovej vrstvy stromov. Montpelierská škola ich zaraďuje do zväzu Ulmeion, Zlatníkova škola do skupiny geobiocénov dubové jaseniny (Fraxini querceta) vyskytujúce sa na pravidelne zaplavovaných miestach a brestové jaseniny (Ulmi fraxineta) vyskytujúce sa na agradačných valoch lemujúcich hlavnú prúdnicu záplavy. Brestové jaseniny sa ďalej delia na topoľovo brestové jaseniny (Ulmi fraxineta-populi) rastúce na vlhkejších miestach a hrabovo brestové jaseniny (Ulmi fraxineta carpini) rastúce na len výnimočne zaplavovaných okrajoch alúvií a predstavujúce prechod k lesom neovplyvňovaným záplavami ani hladinou spodnej vody. Súhrne sa označujú ako tzv. tvrdé luhy.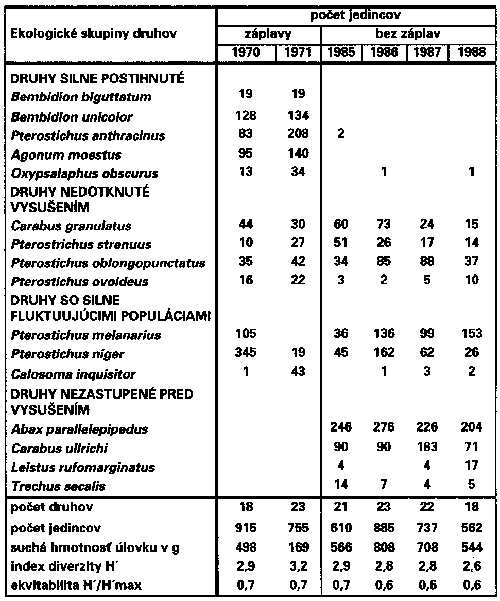
Pravdepodobne už v mladšej dobe kamennej boli lužné lesy vystavené sústredenému tlaku ľudskej populácie. Teplá klíma, plochý reliéf terénu a vysoká produktivita ekosystémov v nivách riek vytvárali priaznivé podmienky pre lov, rybolov, pastvu a poľnohospodárstvo, remeslá i pre zakladanie trvalých sídiel a komunikácií. S rastom ľudskej populácie a rozvojom hospodárstva sa tlak na aluviálne ekosystémy ďalej stupňoval. V niektorých krajinách (Anglicko) vyústil do úplného zničenia lužných lesov už v stredoveku. Inde bol zasadený posledný úder zvyškom prirodzených lužných lesov v minulom storočí pri regulácii riek.
Na Morave, na Slovensku a v Dolnom Rakúsku sa lužné lesy donedávna zachovali v stave a rozsahu, aký inde nemal obdobu. Avšak i tu – na prahu epochy, ktorá pod tlakom ekologických poznatkov začína prehodnocovať antropocentrické myslenie a sústavu hodnôt – velká časť lužných lesov padla v 70. a 80. rokoch za obeť vodným dielam nebývalých rozmerov. Negatívne dopady týchto stavieb a plány na ďalšie stavby na Dunaji a Morave (Hainburg, vodné dielo Nové Mlýny, kanál Dunaj Odra Labe) ohrozujú osud aj posledných zvyškov týchto unikátnych prírodných spoločenstiev. Konflikt ekologických argumentov so záujmami rôznych domácich i zahraničných politických a hospodárskych zoskupení pokúšajúcich sa uskutočniť svoje nové predstavy i staršie zámery preto nekončí. Prevažne technicky alebo humanitne vzdelaná a konzumne zameraná verejnosť sa v zložitej problematike dopadov stavby veľkých vodných diel na krajinu nedokáže orientovať a ťažko môže posudzovať dosah a hierarchiu významnosti predkladaných názorov a argumentov. Vďaka nízkej úrovni ekologického povedomia podlieha technokratickej argumentácii uchyľujúcej sa neraz k vyslovenému zatemňovaniu pojmov, dezinformáciam, sociálnej demagógii a k zneužívaniu národnostného cítenia alebo, azda pod vplyvom zjavných tendencií k stále autoritatívnejším spôsobom vlády, sa o túto problematiku prestáva zaujímať.
Existencia posledných zvyškov relatívne prirodzených aluviálnych ekosystémov na Morave a na Slovensku je preto vážne ohrozená nielen dopadmi už dokončených stavieb, ale aj ďalším plánovitým ničením. Na druhej strane však stále nie je stratená nádej, že časť verejnosti i politikov pochopí závažnosť environmentálnych argumentov a prinúti vládnuce garnitúry k revízii stavebných zámerov i celej koncepcie ekonomiky založenej na pochybnej predstave možnosti neobmedzeného rastu. Nie je preto ani vylúčené, že sa, skôr alebo neskôr, nepristúpi aj k renaturalizácii ekosystémov zničených alebo narušených stavbou vodných diel alebo zbytočnou reguláciou vodných tokov.
Za týchto okolností je potrebné poznať účinky zmien vyvolaných stavbou vodných diel a vodohospodárskymi zásahmi v aluviálnych ekosystémoch. Táto úloha však nie je pri značnej druhovej diverzite aluviálnych ekosystémov zvládnuteľná sledovaním všetkých skupín organizmov. Z časových i metodických dôvodov je naopak výhodné zvoliť niekoľko reprezentatívnych modelových skupín organizmov s rôznym postavením v štruktúre ekosystému, ktoré s potrebnou presnosťou umožňujú prebiehajúce zmeny sledovať v širokej škále stupňov narušenia alebo obnovy ekosystému. Jednou zo skupín, ktoré vďaka neobyčajne bohatej diferencovanosti jednotlivých druhov i spoločenstiev (taxocenóz) vo vzťahu k rôznym ekologickým faktorom umožňujú dôsledky antropických zásahov v lužných lesoch presne sledovať sú prevažne mäsožravé chrobáky z čeľade bystruškovité (Carabidae). Spomedzi asi 550 druhov, ktorými je táto čeľaď zastúpená v strednej Európe, sa asi 150 druhov uplatňuje v taxocenózach bystruškovitých v rôznych typoch lužných lesov. Z tohoto množstva sa na jednej lokalite v prírodných podmienkach vyskytuje súčasne asi 25-35 druhov, iba v najzachovalejších lužných lesoch počet druhov prekračuje 40.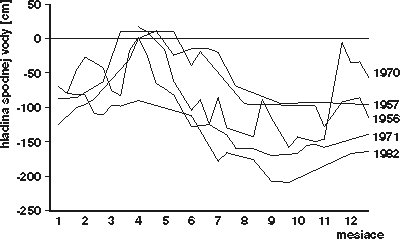
Na lokalitách ovplyvňovaných dlhodobými záplavami s prevahou sedimentačných procesov sa výrazne uplatňujú paludikolné tieňomilné druhy, najmä Carabus granulatus, Bembidion unicolor, Bembidion biguttatum, Oodes helopioides, Pterostichus nigrita, Pterostichus anthracinus, Agonum moestum, Agonum micans, Europhilus (=Agonum) fuliginosus a Platynus (=Agonum) assimilis nahrádzané na niektorých lokalitách na Slovensku vzácnejším druhom Platynus (=Agonum) krynickyi. Na extrémne mokrých s dlhodobo stagnujúcou vodou k nemu pristupuje vzácne i Carabus clathratus.
Na lokalitách s piesčitejším substrátom, zaplavovaných prevažne prúdiacou vodou, naopak prevažuje kombinácia druhov Carabus granulatus, Bembidion femoratum, Asaphidion flavipes, Patrobus atrarufus (=P. excavatus) a Platynus assimilis. Príležitostne, so značnými výkyvmi abundancie medzi jednotlivými rokmi sa v spoločenstvách na takýchto lokalitách uplatňujú Pterostichus niger a Pterostichus melanarius (=vulgaris). Posuv smerom k suchším typom lužných lesov sa prejavuje hlavne úbytkom druhov Bembidion unicolor, Bembidion biguttatum, Oodes helopioides, Pterostichus nigrita, Pterostichus anthracinus. Prirodzene suchší charakter lokalít sa naopak prejavuje postupným pribúdaním niektorých mezohygrofilných druhov s ťažiskom výskytu v lesoch normálneho hygrického radu. Najskôr sa z týchto druhov objavuje Carabus coriaceus, ktorý v suchších variantoch lužných lesov dosahuje obrovskú abundanciu a viaže pozoruhodné množstvo biomasy. K nemu sa postupne pripájajú Carabus ullrichi, Carabus convexus, Pterostichus oblongopunctatus a Abax parallelepipedus (= A. ater) a lokálne Carabus violaceus a vzácne i Carabus scheidleri.
Do plošne rozsiahlych segmentov suchších typov lužných lesov však neprenikajú veľmi typické dominantné druhy listaných lesov normálneho a vysýchavého hygrického radu ako napr. Carabus nemoralis, Carabus hortensis a Abax parallelus. Tieto druhy sa však môžu vyskytovať v úzkych alúviách potokov. Rovnako sa v lužných lesoch výraznejšie neuplatňujú charakteristické ripikolné druhy z rodov Dyschirius, Elaphrus a viaceré druhy z rodov Bembidion, Tachys, Blethisa, Chlaenius a Anisodactylus, Stenolophus a Agonum (s. lat.) viazané na iné typy brehových alebo mokradných biotopov.
Priame pozorovania zmien zloženia taxocenóz bystruškovitých vyvolaných v lužných lesoch poklesom hladiny spodnej vody a vylúčením záplav spoločne so syntézou poznatkov získaných na rôzne narušených lokalitách ukazujú, že uvedené zmeny prebiehajú v zásade tromi možnými smermi.
Prvý smer sa prejavuje postupným poklesom kvantitatívneho zastúpenia a počtu druhov náročných na vlhkosť a ich nahrádzaním druhmi menej náročnými. Uplatňujú sa pri tom len typicky lesné, heliofobné druhy. Výsledkom tohto procesu je spoločenstvo, aké by sme na inom, prirodzene suchšom mieste považovali za celkom prirodzené. Ide tu vlastne len o posuv spoločenstva smerom k suchším hygrickým radom a o proces, aký v medzných úsekoch vlhkostného gradientu prebieha v rôznom rozsahu kontinuálne (a obojsmerne) podľa momentálnych výkyvov vlhkosti a hladiny spodnej vody. Pozoruhodné je, že funkčná štruktúra spoločenstva indikovaná zastúpením rôznych životných foriem sa v takomto prípade mení len málo a to napriek dramatickým zmenám v druhovom zložení taxocenózy.
Takýto proces sme mohli v ostatných 20 rokoch sledovať napr. v Hornom lese pri Lednici na Morave, kde po regulácii Dyje ustali pravidelné jarné záplavy a hladina spodnej vody klesla v ročnom priemere asi o 1 – 1,5 m. Je pritom zrejmé, že takýto druh zmeny, v svojej podstate ešte najprijatelnejší, nemôže nastať hocikde. Jeho predpokladom je totiž zachovanie priestorovej integrity postihnutého porastu a jeho obklopenie alebo susedstvo s prirodzeným lesným spoločenstvom normálneho hygrického radu, ktoré slúži ako zdroj imigrácie i pre málo pohyblivé druhy. Príznačným znakom takto prebiehajúcej zmeny je stále vysoká rozmanitosť spoločenstva.
Druhý smer zmien nastáva v spoločenstvách v niektorých mestských parkoch (napr. Lužánky v Brne alebo Sad J. Kráľa v Bratislave) alebo v niektorých topolových monokulturách. Jeho predpokladom sú málo zmenené vlhkostné pomery a relatívne zachovalý stromový kryt. Na bystruškovité tu pôsobia viac zmeny v bylinnom záraste, charakter hrabanky a opadu, imísie, návštevnosť a pod. Po kvalitatívnej stránke sa pôvodné druhové spektrum viac menej zachováva, no po kvantitatívnej stránke nastávajú značné posuvy v zastúpení jednotlivých druhov. Spravidla sa v takýchto spoločenstvách silne zvýši zastúpenie jedneho alebo dvoch druhov, ku ktorým patrí približne 80-90 % všetkých jedincov. Typická je pre takéto stavy asociácia druhov Platynus assimilis a Patrobus atrarufus. Znakom takéhoto spoločenstva je nízka diverzita pri relatívne vysokej kumulatívnej abundancii. Zdá sa, že takto zmenené spoločenstvo predstavuje v zásade reverzibilný stav. Dokazujú to niektoré vyššie spomínané mestské parky, kde historický prieskum ukazuje, že takéto spoločenstvá môžu predstavovať aj prekvapivo pokročilé štádiá spontánnej, no umelo brzdenej renaturácie.
Tretí smer prebieha za pôsobenia viacerých faktorov. Príznačné je preňho prenikanie veľkého počtu nelesných druhov, zväčša z agrocenóz, výrazným úbytok charakteristických vlhkomilných druhov a nevýrazným zastúpením tolerantnejších lesných druhov normálneho hygrického radu. Samotný počet druhov sa v jednotlivých prípadoch líši. Dokonca aj v takmer zničených zvyškoch lužných lesov môže byť prekvapujúco vysoký. Stretávajú sa tu druhy veľmi rozdielnych ekologických skupín. Jednotlivé druhy sú v konečných štádiích tohoto smeru sukcesie zastúpené zväčša malým počtom jedincov, nepostačujúcim pre nezávislý vývoj populácií. V spoločenstve je vysoký podiel migrantov a zatúlancov z okolitých i vzdialenejších ekosystémov.
Tento typ spoločenstiev je charakteristický pre malé priestorovo značne dezintegrované zvyšky lužných lesov, s narušeným zápojom stromového a krovinného poschodia, odrezané od zdrojov vlhkosti. Je to stav bezprostredne predcházajúci úplnému zániku ekosystémov. Ich príkladom sú izolované zvyšky lužných lesov v okolí bratislavského letiska a Vrakune, ktoré v minulosti lemovali meandre Malého Dunaja.
Na základe pôvodne predpokladaného prietoku v starom koryte Dunaja a vysušenia sústavy ramien bolo možné takéto spoločenstvá oprávnene považovat za predobraz osudu, aký v relatívne krátkom čase po spustení vodného diela Gabčíkovo postihne zvyšné lužné lesy v dotknutej oblasti. Hoci javy pozorované v spoločenstvách bystruškovitých na viacerých lokalitách rok po spustení diela indikujú začiatok sukcesie týmto smerom, zdá sa, že určité zväčšenie prietoku v starom koryte Dunaja a dotovanie zvyškov ramennej sústavy vodou z derivačného kanála a (zatiaľ prisľúbené) simulovanie záplavového režimu vynútené tlakom ochranárskych organizácií degradačný proces spomalí.
Z hľadiska snáh o regeneráciu lužných lesov v oblastiach postihnutých súčasnými vodohospodárskymi úpravami sa však natíska otázka, do akej miery je možná ich regenerácia a aké sú predpoklady jej úspešnosti.
Z hľadiska bystruškovitých je našťastie odpoveď na túto otázku optimistická. Za určitých predpokladov by takáto obnova mohla byť až prekvapujúco rýchla. Na Novomlýnskych nádržiach sme jej začiatok vlastne už mohli sledovať. Keď sa v polovici 80. rokov narušili nečakane silným vlnobitím práve dokončené hrádze II. nádrže, musela byť nádrž počas opráv čiastočne vypustená. Vzniklo tak väčšie množstvo rôzne veľkých ostrovčekov a obnažili sa i pomerne široké ploché pláže. Obnažené plochy začali veľmi rýchlo zarastať pionierskymi spoločenstvami bylín a objavil sa aj rýchlo rastúci nálet topoľov a vŕb, ktorý by pravdepodobne smeroval k vzniku spoločenstiev asociácie Salici-Populetum. Bezprostredné pobrežné pásmo začali rýchlo osidlovať viaceré výrazne hygrofilné druhy bystruškovitých z rodov Bembidion, Dyschyrius a Chlaenius (hlavne Ch. vestitus) a Agonum (s. lat.). Suchšie štrkovité a piesčité miesta ďalej od vody osídlili niektoré vlhkomilnejšie druhy typické (recentne) pre vlhkejšie poľné alebo lúčne stanovisťia, najmä Anisodactylus binotatus, Bembidion lampros a B. properans, Acupalpus spp., Amara spp. a Harpalus rufipes. Druhová rozmanitosť a zloženie vznikajúcich spoločenstiev sa značne približovala relatívne prirodzeným a ustáleným pomerom aluviálnych lúk. Je zrejmé, že zloženie vznikajúcich spoločenstiev by sa začala meniť v závislosti na vývoji drevinnej vegetácie. Ubúdali by svetlomilné druhy a zvyšovalo by sa zastúpenie druhov tieňomilných. Išlo by pravdepodobne o rýchly proces, keďže tieňomilné druhy bystruškovitých tu už boli v malom množstve jedincov prítomné a topoľovo-vŕbové porasty by o 5-10 rokov vytvorili dostatočne hustý zápoj. Sľubný začiatok spontánej obnovy spoločenstiev bystruškovitých prerušilo opätovné napustenie nádrže. Existencia taxocenóz bystruškovitých na vnútornej strane umelých hrádzí krytých záhodzom z lomového kameňa spevneného asfaltom (!) pri plnej hladine nie je možná. Výnimkou sú len ojedninelé konkávne miesta, kde sa usadzuje rôzny organický materiál, na ktorom sa niektoré ripikolné druhy (najčastejšie Agonum ruficorne, na väčších ostrovčekoch náplavu aj Chlaenius vestitus) usadzujú s obdivuhodnou vytrvalosťou. Tieto úseky však netvoria ani zlomok promile vnútorného obvodu hrádzí II. i III. nádrže.
Dočasné čiastočné vypustenie II. nádrže umožnilo priamo v miestnych podmienkach získať prakticky významné poznatky o eventuálnej renaturácii zničených aluviálnych ekosystémov. Obnova taxocenóz bystruškovitých by s veľkou pravdepodobnosťou začala okamžite po vypustení nádrží. Už v jej iniciálnych štádiách by sa tu objavili druhy, ktoré vytvárajú prirodzené spoločenstvá tak v lužných lesoch ako aj v trstinách a na iných mokradných biotopoch, ktoré by na mieste nádrže bolo žiadúce obnoviť. Na priebeh sukcesie by priaznivo pôsobili aj spoločenstvá v priľahlých agrocenózach, kde niektoré druhy uplatňujúce sa v aluviálnych spoločenstvách vcelku úspešne prežívajú. Niektoré z ních, napr. Pterostichus melanarius, bývajú tak v lužných lesoch ako aj na poliach dokonca veľmi početne zastúpené. Táto zdanlivo prekvapujúca skutočnosť vyplýva z hypotetického, ale v ostatnom čase čoraz širšie prijímaného predpokladu, že značná časť druhov bystruškovitých zastúpená v poľných a lúčnych ekosystémoch pôvodne obývala suchšie časti alúvií porastené vyššou bylinnou vegetáciou. Vďaka určitým zmenám ekologických nárokov niektorých druhov predstavujú dnes polia dokonca akési „refúgium“ alebo „náhradný biotop“ i pre pôvodne tak vyhranene mokradný druh ako Drypta dentata. Regenerácia společenstiev bystruškovitých v porastoch trstiny alebo ostric by bola podporovaná imigráciou niektorých druhov z polí ešte viac.
I v silne devastovanom prostredí okolo Novomlýnskych nádrží (a ďalších podobných stavieb) ostávajú ostrovčeky poloprirodzenej vegetácie, v ktorých prežívajú aspoň najhojnejšie a najtoleratnejšie druhy (napr. Oxypselaphus (=Agonum) obscurus, Platynus assimilis, Agonum moestum) typické pre pôvodné, veľkoplošne zničené lužné ekosystémy. Rovnaký význam majú napriek silnému tlaku divokých rekreantov i malé úseky brehov nespevnené asfaltom alebo betónom.
Najvýznamnejším argumentom pre úspešnosť obnovy ekosystémov zničených stavbou Novomlýnskych nádrží je existencia komplexu porastov okolo Křivého jazera pri Nových Mlýnoch a pod obcou Iváň, kde sa až do súčasnosti udržali spoločenstvá bystruškovitých, ktoré sa svojím zložením podobajú spoločenstvám, aké v tejto oblasti žili pred začiatkom stavby nádrží a reguláciou Dyje pod nádržami. Tieto porasty v prípade rozumne riadenej renaturácie možu slúžiť ako významná rezerva pre spontánnu imigráciu alebo priamu introdukciu druhov na renaturalizované územie. Podobná situácia je i v oblasti Gabčíkova, hoci rozsah a intenzita devastácie krajiny sú tam oveľa väčšie.
Pre úspešnosť budúcich pokusov o obnovu narušených alebo úplne zničených lužných lesov však hovoria aj niektoré všeobecné poznatky. Jedným z veľmi dôležitých adaptačných mechanizmov vlhkomilných druhov žijúcich na brehoch vôd a v alúviách je veľká pohyblivosť a schopnosť migrácie. Pri náhlych záplavách alebo zmenách vlhkosti preto môžu rýchlo opustiť zničené alebo prechodne nevhodné stanovišťa a presídliť na trvale alebo dočasne vhodnejšie miesta. Táto schopnosť má mimoriadny význam najmä v neobyčajne dynamicky sa meniacich alúviách divočiacich tokov. Vysoká migračná schopnosť umožňuje vyslovene hygrofilným a ripikolným druhom úspešne obsadzovať dokonca i brehy efemérnych jazierok roztrúsených v rozsiahlych púštiach. Pozoruhodný príklad takýchto druhov predstavuje v našej faune bystruška Carabus clathratus. V strednej Európe je veľmi lokalizovaným druhom viazaným na lokality s dlhodobo stagnujúcou vodou (napr. Jurský Šúr pri Bratislave alebo niektoré miesta na Morave). V púštiach Strednej Ázie (Mujunkum, Kyzylkum) je však pri obsadzovaní brehov dočasných jazierok rovnako úspešná ako napr. behúnik Pterostichus nigrita, ktorého nájdeme takmer pri každej mokradi. Vysokú schopnosť migrácie a propagácie hygrofilných druhov bystruškovitých dokazuje i zloženie taxocenóz, alebo presnejšie povedané dočasných agregácií, bystruškovitých na rozličných lokalitách v strede veľkomiest. Medzi celkovo veľmi malým počtom jedincov bystruškovitých, ktoré sem rôznymi cestami prenikajú, vlhkomilné druhy výrazne prevažujú. V asfaltovo-betónovej púšti našich miest je to na prvý pohľad paradox. Veľká propagačná schopnosť vlhkomilných druhov vyniká aj pri porovnaní veľkosti areálov geografického rozšírenia druhov s rôznymi ekologickými nárokmi. Medzi vlhkomilnými druhmi je oveľa väčší podiel druhov s veľkými areálmi rozšírenia ako medzi druhmi uprednostňujúcimi iné typy stanovíšť.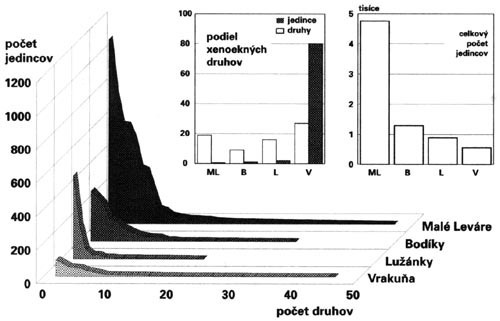
Existujú aj príklady rýchleho spontánneho vzniku neobyčajne bohatých aluviálnych alebo brehových spoločenstiev bystruškovitých v relatívne málo priaznivých podmienkach. Jedným z ních je spoločenstvo na brehoch Jašterkového jazierka v Slovenskom krase. Toto jazierko vzniklo upchatím dna plochého závrtu, v ktorom bolo ešte pred vojnou pole. V súčasnosti bystruškovité na jeho brehoch tvoria zriedka vídaný bohatý súbor ekologicky vyhranených a neraz aj pomerne vzácnych druhov napriek tomu, že najbližšimi potenciálnymi imigračnými zdrojmi sú len nevyhranené ekotonálne spoločenstvá vo vzdialenosti asi 2,5 km. Pozoruhodný prípad spontánnej obnovy lužného spoločenstva bystruškovitých nájdeme v parku Lužánky v Brne. Toto spoločenstvo vzniklo samovoľne v parku založenom r. 1780 na mieste úplného bezlesia. Vďaka silnému antropickému tlaku síce sukcesne stagnuje približne na stupni, aký je charakteristický pre narušené topoľové monokultúry v alúviu Dunaja. Vyskytujú sa tu však všetky druhy, ktoré nachádzame aj v prirodzených lužných lesoch. Preto za priaznivých podmienok má toto spoločenstvo vnútorné predpoklady svoju prirodzenú štruktúru úplne obnoviť.
Doterajšie poznatky ukazujú, že spoločenstvá bystruškovitých by sa na renaturalizovaných územiach devastovaných stavbou vodných diel rýchlo obnovovali. Bez väčších ľudských zásahov by pravdepodobne už za 10-15 rokov dosiahli značne pokročilý stupeň sukcesie. Umelá reintrodukcia niektorých menej pohyblivých druhov by jej priebeh ešte urýchlila.
Samozrejme obnova taxocenóz jednotlivých skupín organizmov, resp. hmyzu nebude prebiehať rovnako rýchlym tempom ako u bystruškovitých. Rýchlosť ich obnovy bude v rozhodujúcej miere závisieť na rýchlosti, s akou budú jednotlivé druhy rastlín, najmä drevín, dospievať do štádia potrebného pre vývin toho-ktorého druhu hmyzu. Nosáčikovité (Curculionidae) alebo liskavkovité (Chrysomelidae) či motýle začnú vytvárať svoje spoločenstvá, keď sa uchytia živné rastliny niektorých druhov. Niektoré druhy fúzačov (Cerambycidae), napr. fúzač obrovský (Cerambyx cerdo), tu nájdu vhodné podmienky až po 80 – 100 rokoch. Pre vývoj niektorých vysoko špecializovaných kováčikov (Elateridae) žijúcich v dutinách stromov nastanú podmienky ešte neskôr. Pri vzácnosti niektorých druhov hmyzu nemôžeme očakávať, že renaturalizované ekosystémy lužných lesov sa vrátia do úplne pôvodného stavu skôr ako za 150-200 rokov. To je však hraničná doba pre najnáročnejšie druhy hmyzu. Reálne môžeme predpokladať, že značne pokročilý stupeň sukcesie dosiahne väčšina taxocenóz živočíchov v tzv. tvrdých luhoch za 60 až 80 rokov a v mäkkých za 20 až 30 rokov, v trávnatých alebo trstinových spoločenstvách možno za ešte kratší čas.
Súčasné ekologické poznatky umožňujú vytvoriť veľmi konkrétne predstavy o renaturalizácii aluviálnych spoločenstiev a o spôsoboch jej usmerňovania. Základným predpokladom renaturálizácie zničených aluviálnych ekosystémov je však obnova vodného režimu, ktorý by existenciu obnovených ekosystémov umožňoval bez dalších ľudských zásahov. Ale aj tento predpoklad je splniteľný. A tak sa zdá, že najväčšou prekážkou obnovy zničených lužných ekosystémov budú záujmy rôznych hospodárskych a politických zoskupení, pristupujúcich k rozhodovaniu o veľkých vodných dielach a životnom prostredí z pozíc vyhraneného skupinového egoizmu a myšlienkovou a vedomostnou výbavou vyjadrenou heslom:
„BUDUJEME NÁRODNÝ PARK PRE ĽUDÍ A NIE PRE KOMÁROV“
O autorovi
Zbyšek Šustek
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
