










Řiť i čelo – jedno tělo aneb bříškem vzhůru
Hlava a ocas byly v zmíněném článku Pavly Loucké pojaty jako protilehlé konce těla, jeden důležitý, druhý nevýznamný. V tom duchu se odvíjely různé významové souvislosti v češtině, uplatnění hlavy i ocasu v ustálených slovních spojeních, v názvosloví a jinde. Živé organizmy ovšem nemají jenom „začátek“ a „konec“, ale i další dimenze. A toto všechno vzniká podle určitého, předem daného „programu“.
Většina živočišných kmenů patří do velké skupiny organizmů dvoustranně souměrných – Bilateralia. Ano, patříme sem i my, i když nás J. David na svých fotografiích (Vesmír 76, 58, 1997/1) přesvědčuje, že náš obličej zdaleka není zrcadlením dvou identických půlek, nemluvě už o značně asymetrickém rozložení našich útrob; patří sem i ostnokožci (hvězdice, ježovky) se svou pětičetnou symetrií, a dokonce spirálovitě všelijak pokroucení a naprosto asymetričtí plži. Laik se bude divit podobně, jako když se dovídá, že hadi a velryby patří mezi čtyřnožce. A přesto má toto třídění logiku – když ne v dospělosti, tak alespoň v raných stadiích vývoje lze na těle rozeznat tři osy souměrnosti: předozadní (od hlavy k ocasu), dorzoventrální (od hřbetní strany k břišní) a pravolevou (ano, i u hvězdic). Podobně lze argumentovat v případě chybějících končetin hadů a velryb.
Skupina Bilateralia se opět člení na dvě velké skupiny: na prvoústé (Protostomia), kam ze známějších skupin patří členovci a měkkýši, a druhoústé (Deuterostomia), kam patří i zmínění ostnokožci a obratlovci. Abychom pochopili, proč toto dělení, musíme nejdříve k raným stadiím utváření tělní stavby. Ze školy si možná pamatujeme, že rýhováním oplozeného zárodku – zygoty – vznikne shluk buněk – morula. V ní se postupně utvoří dutina, buňky se uspořádají na povrchu do jedné vrstvy a vznikne blastula. Tento „balón“ se začne na jednom místě vchlipovat a vzniká gastrula (obr. obrázek). 
Gastrulace je nejdůležitějším obdobím utváření tělní stavby. Dutina vchlípeniny je dutinou příští trávicí soustavy; průchodnost se zajistí např. tak, že tam, kde se vchlípenina nakonec dotkne protější strany koule, se prorazí druhý otvor. Místo vchlipování se nazývá prvoústa a samotná gastrulace je kombinací vtahování či vtlačování materiálu dovnitř (jako u propíchnutého míče), ale zejména vcestováním buněk z vnějšího povrchu koule kolem rtů prvoúst dovnitř. Rty jsou možná klíčovým řídícím centrem celého morfogenetického procesu: procházející buňky zde dostávají důležité informace o svých časových a prostorových souřadnicích, a z toho plynoucích „kompetencích“ k dalšímu vývoji. Druhým řídícím momentem je indukce – vzájemné ovlivňování vnitřní vrstvy – endodermu a vnější vrstvy – ektodermu. V souhře obou dějů je zaplňována dutina mezi ektodermem a endodermem buď vychlipováním váčků z obou vrstev (tak vzniká nervová soustava, játra, plíce), nebo vycestováním skupin buněk, které dají vznik mezodermu a z něho odvozeným orgánům, např. svalům, ledvinám, srdci, pohlavním orgánům. (Není třeba zdůrazňovat, že nově vzniklé mezodermální orgány se aktivně zapojují do dalších indukčních procesů.)

Takhle to ovšem probíhat nemuselo. Druhoústí nemají vzhledem k prvoústým hlavu na neslušném místě těla. Předozadní osa u prvoústých i druhoústých je orientována shodně, a dokonce její jednotlivé úseky jsou determinovány homologickými sadami genů, z nichž nejpozoruhodnější a nejlépe prozkoumané jsou tzv. homeotické geny (Vesmír 74, 276, 1995/5; 75, 7, 1996/1 a článek F. Sládečka v tomto čísle). Homologie jde ještě dál. Jednotlivé struktury mozku jsou – z hlediska raných stadií vývoje a sítě regulací – také homologickými útvary.[2,3] Tedy o homologii se přestává mluvit na úrovni struktur, ale mluví se o ní na úrovni procesů, které ke strukturám vedou, což určitě příliš nepotěší taxonomy[3]. Totéž platí pro deriváty mozku – oči (Vesmír 74, 495, 1995/9 a článek F. Sládečka na v tomto čísle, 75, 92, 1997/2 ).
Zkusme si vzít na pomoc druhou osu symetrie – dorzoventrální. Už v polovině minulého století si povšiml Geoffroy Saint–Hillaire
(viz obr. 1
obrázek v článku F. Sládečka na s. 92, 75, 92, 1997/2 ), že prvoústí se od druhoústých liší postavením vnitřních orgánů: prvoústí mají nervovou soustavu na břišní straně, srdce na straně hřbetní, u druhoústých je to právě obráceně; a týká se to řady dalších orgánů. A najednou se ukazuje, že regulační signály pro určení této osy jsou u obou skupin identické, jenom jsou – otočeny vůči sobě. Určuje-li tedy u octomilky (Drosophila, moucha, tj. tvor prvoústý) produkt genu A vznik hřbetních struktur a produkt genu B struktury břišní, pak jejich homology A’, B’ u obratlovců budou působit v obráceném registru (obr. obrázek, podrobněji v zmíněném článku F. Sládečka).[5,8] 
{kind=link}
U octomilky mutace genu B (dorsal) opravdu způsobí, že po celém obvodu těla se vyvinou jen hřbetní struktury. Ne vždy to ovšem vychází tak jednoznačně, protože pod symboly A, B se skrývají celé baterie regulačních genů... ovšemže opět homologických mezi oběma velkými skupinami živočichů. A homologie jde i na nižší úroveň: např. kdo by byl předpokládal ještě před pár lety, že indukce vývoje srdce je totožná u mouchy i u člověka!
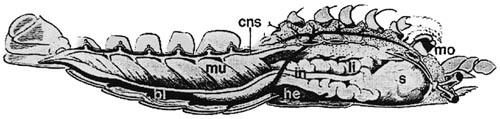
S touto znalostí to zkusme jinak. Trávicí roura nemusí u prvoústých vznikat proražením řitního otvoru, může vzniknout i srůstem „koutků“ prvoúst gastruly (obr. obrázek).[4]  Pak už zbývá jen jedno. Druhoústí si z neznámých příčin vytvořili nová ústa na temeni a poté se otočili „bříškem vzhůru“ tak, aby se ústní otvor nacházel na „správné“ straně. Dokonce snad existuje jakýsi přechodný stav dochovaný u druhoústého kmene Enteropneusta (žaludovci, obr. obrázek).[7]
Pak už zbývá jen jedno. Druhoústí si z neznámých příčin vytvořili nová ústa na temeni a poté se otočili „bříškem vzhůru“ tak, aby se ústní otvor nacházel na „správné“ straně. Dokonce snad existuje jakýsi přechodný stav dochovaný u druhoústého kmene Enteropneusta (žaludovci, obr. obrázek).[7] 
Když už je tento příspěvek reakcí na článek P. Loucké, tak ještě k hlavě a ocásku. Podle jedné teorie je ocas pokračováním gastrulace. Je-li vnitřní organizace gastruly u konce, začne se materiál proudící kolem horního rtu prvoúst městnat na okraji, takže vznikne jakýsi pupen, a protože nemůže dovnitř, pokračuje v růstu směrem ven.
Mnohem zajímavější je to s hlavou. Už morfologové si všimli, že „primitivní“ formy vývojových linií nemají hlavu v pravém slova smyslu – srovnejte dvojici žížala – moucha nebo kopinatec – ryba (tím nechci tvrdit, že první z dvojice je předkem druhého). Tedy – hlava jako něco nového, druhotného, co vzniklo v evoluci nezávisle u obratlovců i u členovců[9]. Molekulární biologie zpočátku této teorii nahrávala – definice tělních částí na předozadní ose pomocí homeotických genů nezačíná od prvního koncového článku, ale až na úrovni čelistních článků u hmyzu a na úrovni zadního mozku u obratlovců. Mozek a hlava kolem něho jakoby vyrašily až později, u obou skupin nezávisle, a jejich vznik je regulován jinak než homeotickými geny. Posléze se však zjišťuje,[2] že tytéž regulační obvody determinují vývoj mozku u obou skupin, a na základě tohoto poznání lze dokonce provést anatomickou homologizaci mezi některými strukturami lidského a mušího mozku.
Takže jak to bylo? Jedna zajímavá teorie [10] nabízí tento scénář: Někdy v kambriu se u jedné skupiny drobných mnohobuněčných organizmů podobných larvám dnešních kroužkovců nebo ostnokožců objevila schopnost generovat nové, často k přežití naprosto nepotřebné článkované výrůstky. Protože samotné přežití zajišťovaly ony původní organizmy ještě než jim tyto novotvary na těle vyrostly, nebylo třeba se příliš omezovat vůči přírodnímu výběru a došlo ke „kambrické explozi“, onomu neuvěřitelnému třeštění tvarů a forem, se kterým si dodnes evolucionisté lámou hlavu. (Vesmír 70, 305, 1991/6; tam už je ale líčena pokročilejší fáze tohoto vývoje.) Některé z těchto forem pak začaly žít na vlastní pěst, vzaly na sebe i břemeno rozmnožování a daly tak vzniknout všem dnes žijícím živočišným kmenům skupiny Bilateralia. A snad při všech těch excesech a tvarových experimentech některé zkusily, co to udělá, když si prostrčí ústa na temeno a posléze se otočí bříškem vzhůru. A díky nim jsme tady.
Anebo: Je možné pojmout homologie na úrovni genů (sekvence), regulačních drah (např. homeotické geny) a struktur (křídlo ptáka, křídlo netopýra) do jednoho uceleného a bezrozporného systému? Někteří autoři (např. [3]) to popírají. Nebude to spíš tak, že některé regulační moduly budou používány vždy znovu a znovu v nejrůznějších kontextech? Analogie trochu přitažená za vlasy, ale když už jednou byly zavedeny metrické šrouby, tak najdeme šroub M6 v ponorce i proudovém letadle, a když byl vynalezen karburátor, tak se vyskytne ve škodovce i v motorovém člunu... Víme už, že na úrovni proteinů si evoluce hrála s moduly: má-li třeba enzym vázat nějaký ligand, např. ATP či ionty vápníku, nepostupuje evoluce „vymýšlením“ modulu vázajícího ATP či vápník, pokud takový modul už existuje a lze jej k molekule enzymu prostě připojit. Půjde to zobecnit i na struktury vyššího řádu? Když je potřeba rozčlenit nějaký prostor, hodí se sada homeotických genů, ať je tím prostorem celé embryo, končetina nebo třeba ledvina. Když je potřeba definovat ostré rozhraní mezi dvěma strukturami (hmyzí články, hmyzí křídlo, oddíly mozku obratlovců atd.), opakovaně se objeví modul 3–4 homologických regulačních genů. Když je potřeba postavit oko, mozek, srdce...? Zdá se, že vstupujeme do velmi zajímavé doby.
Literatura
[1] Gilbert S. F.: Developmental biology, 4th edn. Sinauer; 1994[2] Arendt D., Nubler-Jung K.: Common ground plans in early brain development in mice and flies. BioEssays 18, 255–259, 1996
[3] Bolker J., Raff R. A.: Developmental genetics and traditional homology. BioEssays 18, 489–494, 1996
[4] Lacalli T.: Dorsoventral axis inversion: a phylogenetic perspective. BioEssays 18, 251–254, 1996
[5] Jones C. M., Smith J. C.: Inductive signals: revolving vertebrates. Curr. Biol. 5, 574–576, 1995
[6] Tam P. P. L., Quinlan G. A.: Mapping vertebrate embryos. Curr. Biol. 6, 104–106, 1996
[7] Nubler-Jung K., Arendt D.: Enteropneusts and chordate evolution. Curr. Biol. 6, 352–353, 1996
[8] De Robertis E. M., Sakai Y.: A common plan for dorsoventral patterning in Bilateria. Nature 380, 37–40, 1996
[9] Gans C., Northcutt R. G.: Neural crest and the origin of vertebrates: a new head. Science 220, 268–274, 1983
[10] Davidson E. H., Peterson K. J., Cameron R. A.: Origin of bilaterian body plans: Evolution of developmental regulatory mechanisms. Science 270, 1319–1325, 1995
O autorovi
Anton Markoš
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
