










Subsaharské dcery pramatky Evy
 Nedávno vyšel český překlad úspěšné knihy britského genetika Bryana Sykese Sedm dcer Eviných. 1) Autor líčí své pokusy izolovat DNA z prastarých koster, rozebírá genetické souvislosti osídlování tichomořských ostrovů a přibližuje postupy, pomocí nichž můžeme z poznatků o současné molekulární rozmanitosti mitochondriální DNA odhalovat historii dnešních populací. Knihu uzavírají úvahy o přírodním prostředí a osudech hypotetických „sedmi dcer Eviných“, od nichž by mohli odvozovat svůj původ současní Evropané. Těchto „sedm dcer“ však zdaleka nepředstavuje jediné potomky naší východoafrické pramatky, větší část jejích dcer zůstala v Africe. 2)
Nedávno vyšel český překlad úspěšné knihy britského genetika Bryana Sykese Sedm dcer Eviných. 1) Autor líčí své pokusy izolovat DNA z prastarých koster, rozebírá genetické souvislosti osídlování tichomořských ostrovů a přibližuje postupy, pomocí nichž můžeme z poznatků o současné molekulární rozmanitosti mitochondriální DNA odhalovat historii dnešních populací. Knihu uzavírají úvahy o přírodním prostředí a osudech hypotetických „sedmi dcer Eviných“, od nichž by mohli odvozovat svůj původ současní Evropané. Těchto „sedm dcer“ však zdaleka nepředstavuje jediné potomky naší východoafrické pramatky, větší část jejích dcer zůstala v Africe. 2)
 Modrým (nebo černým v obrázcích) textem jsou vyznačeny haploskupiny, o nichž se v článku pouze píše, červeným pak haploskupiny, které autor při výzkumu detekoval.
Modrým (nebo černým v obrázcích) textem jsou vyznačeny haploskupiny, o nichž se v článku pouze píše, červeným pak haploskupiny, které autor při výzkumu detekoval.
Kouzla s mitochondriální DNA, a nejen s ní
Vždycky jsem trochu záviděl zoologům a botanikům, jak relativně snadno si opatřují výzkumný materiál. Ti, kteří se zabývají populační genetikou člověka, musí obvykle škemrat u vrat nemocnic či transfuzních stanic, shánět dobrozdání etických komisí a dalších institucí. Musí putovat do odlehlých regionů, protože dostupné městské populace jsou nepřehledně promíchány. Se svými „vzorky“ musí hovořit, vysvětlovat jim, přesvědčovat je, potýkat se s jazykovými problémy. Potřebují místního asistenta – rodilého mluvčího, který lidem vysvětlí, o co tomu bílému podivínovi vlastně jde, a pomůže vybrat nepříbuzné dobrovolníky. Bez jeho pomoci je výzkum genetické pestrosti odlehlých afrických oblastí téměř nemyslitelný.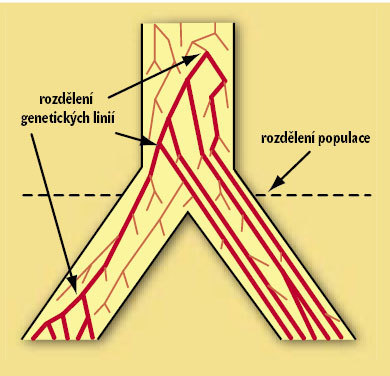 Genetické složení každé populace je výsledkem pravěkých demografických událostí, o jejichž rekonstrukci se pokoušíme. Pro tato studia se ujal termín archeogenetika. Zahrnuje také analýzy pravěké DNA izolované z kosterních nálezů, ale zvláštní postavení v archeogenetickém výzkumu současných populací hrají nerekombinantní úseky DNA, tedy mitochondriální DNA (mtDNA) děděné po matce (matrilineárně) a větší úsek chromozomu Y (NRY), který se dědí pouze z otců na syny. Mluvíme obvykle o haplotypech, 3) které generacemi procházejí pouze skrze jedno pohlaví a jejich rozmanitost se zvyšuje jen mutacemi.
Genetické složení každé populace je výsledkem pravěkých demografických událostí, o jejichž rekonstrukci se pokoušíme. Pro tato studia se ujal termín archeogenetika. Zahrnuje také analýzy pravěké DNA izolované z kosterních nálezů, ale zvláštní postavení v archeogenetickém výzkumu současných populací hrají nerekombinantní úseky DNA, tedy mitochondriální DNA (mtDNA) děděné po matce (matrilineárně) a větší úsek chromozomu Y (NRY), který se dědí pouze z otců na syny. Mluvíme obvykle o haplotypech, 3) které generacemi procházejí pouze skrze jedno pohlaví a jejich rozmanitost se zvyšuje jen mutacemi.
 Mutace mtDNA nejsou nic jiného než odchylky od Cambridžské referenční sekvence, která byla prvně publikována r. 1981. Většinou jde o náhrady jednoho nukleotidu za druhý, častější jsou tranzice, 4) méně časté transverze. 5) Mutace mtDNA označujeme číslem, které udává pořadí odpovídajícího nukleotidu v referenční sekvenci. Můžeme porovnávat různé haplotypové sekvence a stopovat tak mitochondriálního předka.
Mutace mtDNA nejsou nic jiného než odchylky od Cambridžské referenční sekvence, která byla prvně publikována r. 1981. Většinou jde o náhrady jednoho nukleotidu za druhý, častější jsou tranzice, 4) méně časté transverze. 5) Mutace mtDNA označujeme číslem, které udává pořadí odpovídajícího nukleotidu v referenční sekvenci. Můžeme porovnávat různé haplotypové sekvence a stopovat tak mitochondriálního předka.
 V případě NRY jsou mutace rozptýleny v celém dlouhém úseku, kdežto u mtDNA jde obvykle jen o úsek krátký – první hypervariabilní segment (HVS-I) čítající 360 párů bází (od pozice 16 030 do 16 390), který patří do nekódující oblasti – D-smyčky. Údaje o mutacích v HVS-I (mutační motiv) mohou doplnit informace z druhého hypervariabilního segmentu (HVS-II) a z kódující části molekuly mtDNA. Obecně platí, že čím je haplotyp mladší (méně odlišný), tím více informací potřebujeme k tomu, abychom jej zařadili do jedné z mnoha známých haploskupin, tedy sady fylogeneticky si příbuzných haplotypů.
V případě NRY jsou mutace rozptýleny v celém dlouhém úseku, kdežto u mtDNA jde obvykle jen o úsek krátký – první hypervariabilní segment (HVS-I) čítající 360 párů bází (od pozice 16 030 do 16 390), který patří do nekódující oblasti – D-smyčky. Údaje o mutacích v HVS-I (mutační motiv) mohou doplnit informace z druhého hypervariabilního segmentu (HVS-II) a z kódující části molekuly mtDNA. Obecně platí, že čím je haplotyp mladší (méně odlišný), tím více informací potřebujeme k tomu, abychom jej zařadili do jedné z mnoha známých haploskupin, tedy sady fylogeneticky si příbuzných haplotypů.
V celosvětovém měřítku je mtDNA jednoznačně nejrozmanitější u subsaharských populací, zejména pak ve východní Africe. Proto máme za to, že zde kdysi žila naše „mitochondriální pramatka“ a stačí nám i poměrně málo informací k tomu, abychom příslušný haplotyp z fylogenetického hlediska správně zařadili. Stanovením mutační rychlosti daného úseku molekuly mtDNA (odhadu, jak rychle tyto molekulární hodiny tikají) lze usuzovat, kdy asi tato žena žila. Dnes mluvíme o době zhruba před 150–200 tisíci lety, a tím vylučujeme možnost, že by se na rozmanitosti mtDNA současného lidstva nějak podílel člověk vzpřímený (Homo erectus), který osídlil Asii a Evropu asi před milionem let. Dokonce i výzkum mtDNA z kostí neandertálců a anatomicky moderních kromaňonců ukazuje, že se tyto dvě skupiny nejspíše vůbec nemísily.

 Někteří badatelé dávají současné poznatky do souvislosti s tezí o monofyletickém původu anatomicky moderního člověka (Homo sapiens sapiens), což je však poněkud zavádějící. Odhad období, kdy došlo k prvnímu rozrůznění mtDNA, se totiž nevztahuje k ničemu jinému než právě jen k tomu úseku mtDNA, který posloužil výpočtu. Ostatní části genomu, které ovlivňují fyziologické vlastnosti či stavbu a tvar těla, jsou na onom úseku mtDNA obvykle nezávislé (obrázek 2).
Někteří badatelé dávají současné poznatky do souvislosti s tezí o monofyletickém původu anatomicky moderního člověka (Homo sapiens sapiens), což je však poněkud zavádějící. Odhad období, kdy došlo k prvnímu rozrůznění mtDNA, se totiž nevztahuje k ničemu jinému než právě jen k tomu úseku mtDNA, který posloužil výpočtu. Ostatní části genomu, které ovlivňují fyziologické vlastnosti či stavbu a tvar těla, jsou na onom úseku mtDNA obvykle nezávislé (obrázek 2).
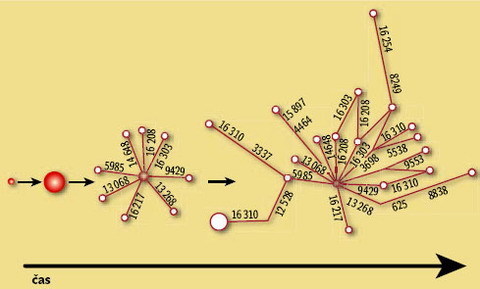
Řada studií ukazuje, že k rozrůznění určité linie mtDNA může dojít jen při expanzi početně nevýznamné populace. Představme si ženu (sekvenci mtDNA), jejíž potomci díky reprodukčnímu úspěchu (při zlepšení klimatických podmínek, osídlení nového teritoria s bohatšími potravními zdroji apod.) početně vzrostou (viz obrázek 4). Během času někteří z nich přirozeně zmutují a nepatrnou změnou v pozici jednoho nukleotidu se odliší od své mateřské mtDNA. Mutace jsou v podstatě náhodné (většina evolučně nestabilních pozic je dnes již známa), proto po čase vznikne (za nezměněných podmínek populační expanze) hvězdicovité uspořádání fylogenetické sítě tohoto celého matrilineárního příbuzenstva. Jak haploskupina stárne, původní typ se postupně stává méně a méně početným. 6)
Dnešní scénář vzniku subsaharských mtDNA
V subsaharské Africe, kde „na to byl čas“, se mtDNA rozrůznila mnohem víc než kdekoli jinde na světě. Najdeme tu ne sedm, ale hned několik desítek dcer, vnuček a pravnuček naší africké pramatky, jakýchsi našich „tet“ a „sestřenic“. Některé se vyskytují pouze u konkrétních etnických skupin, což je však spíše ojedinělé. 7) K nejstarší demograficky významné události, kterou jsme schopni odhalit, došlo v subsaharské Africe zhruba před 150–200 tisíci lety. Tehdy vznikla haploskupina nazvaná L1, od níž se oddělily L2, L3, L5 a L6 (obrázek 5). Z okruhu L3 se odštěpily M a N, jejichž nositelé asi před 60–80 tisíci lety opustili Afriku a nejspíše úžinou Báb el-Mandab osídlili Arabský poloostrov, a v poměrně krátké době také indický subkontinent (M) a západní Euroasii (N). Některé výzkumy připouštějí, že M a N mohly vzniknout v Asii, přičemž do Afriky je mohla zanést jedna ze zpětných migrací během posledních 20 tisíc let. Nelze vyloučit ani to, že haploskupina M vznikla až v Indii. Ať tak či onak, jsou všechny současné neafrické linie mtDNA potomky poměrně nevýznamné části L3. V subsaharské Africe vznikly haploskupiny, které dnes označujeme L0 až L6. Na obrázek 5 uvádíme nejdůležitější z jejich potomků (dcer).
K nejstarší demograficky významné události, kterou jsme schopni odhalit, došlo v subsaharské Africe zhruba před 150–200 tisíci lety. Tehdy vznikla haploskupina nazvaná L1, od níž se oddělily L2, L3, L5 a L6 (obrázek 5). Z okruhu L3 se odštěpily M a N, jejichž nositelé asi před 60–80 tisíci lety opustili Afriku a nejspíše úžinou Báb el-Mandab osídlili Arabský poloostrov, a v poměrně krátké době také indický subkontinent (M) a západní Euroasii (N). Některé výzkumy připouštějí, že M a N mohly vzniknout v Asii, přičemž do Afriky je mohla zanést jedna ze zpětných migrací během posledních 20 tisíc let. Nelze vyloučit ani to, že haploskupina M vznikla až v Indii. Ať tak či onak, jsou všechny současné neafrické linie mtDNA potomky poměrně nevýznamné části L3. V subsaharské Africe vznikly haploskupiny, které dnes označujeme L0 až L6. Na obrázek 5 uvádíme nejdůležitější z jejich potomků (dcer).
Haploskupina L0d, jejíž vznik spadá do doby před 50 tisíci lety, se vyskytuje především u Khoisanů v jižní Africe. Nalezneme ji však také v jihovýchodní části kontinentu a ojediněle v Africe východní – dosud byla zjištěna pouze u jedince z etnika Turkana. Je třeba zmínit teorii předpokládající, že Khoisané žili dříve na celém území jižní, jihovýchodní a východní Afriky. Tito lidé, živící se ještě dnes převážně lovem a sběrem, používají v řeči mlaskavky, jež se vyskytují i v jazycích současných východoafrických Hadzů a Sandawů. Má se za to, že mlaskavky jsou jakousi jazykovou stopou ukazující původní rozšíření khoisanské populace. L0d však u „mlaskajících“ Hadzů ani Sandawů dosud doložena nebyla. Dokládá to, jak komplikovaný a často spíše nezávislý je vývoj jazykových (získaných) a biologických (vrozených) struktur.
 Subsaharské haploskupiny mtDNA jsou velmi rozmanité a jen omezený počet vznikl v holocenním období (za posledních 10 tisíc let). Není divu, vždyť v době, kdy Afričané přijali zemědělskou technologii, byli již natolik početní, že přirozeně vznikající mutace, které by mohly další haploskupiny vytvořit, většinou neuspěly ve velké konkurenci již existujících typů mtDNA. Subsaharské zemědělství, využívající zejména čirok (Sorghum bicolor) a dochan (Pennisetum glaucum), je ostatně záležitostí posledních zhruba dvou tisíc let. Do té doby žila většina Afričanů z přebohaté nabídky přirozených potravních zdrojů, které jejich kontinent nabízel celoročně. Jedna z tezí o súdánském pásmu v holocenním období mluví dokonce o „rajské zahradě“.
Subsaharské haploskupiny mtDNA jsou velmi rozmanité a jen omezený počet vznikl v holocenním období (za posledních 10 tisíc let). Není divu, vždyť v době, kdy Afričané přijali zemědělskou technologii, byli již natolik početní, že přirozeně vznikající mutace, které by mohly další haploskupiny vytvořit, většinou neuspěly ve velké konkurenci již existujících typů mtDNA. Subsaharské zemědělství, využívající zejména čirok (Sorghum bicolor) a dochan (Pennisetum glaucum), je ostatně záležitostí posledních zhruba dvou tisíc let. Do té doby žila většina Afričanů z přebohaté nabídky přirozených potravních zdrojů, které jejich kontinent nabízel celoročně. Jedna z tezí o súdánském pásmu v holocenním období mluví dokonce o „rajské zahradě“.
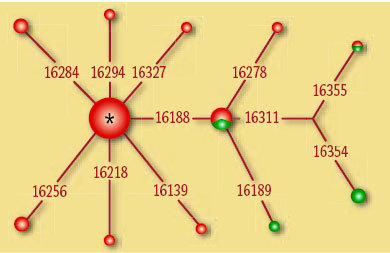 Současná představa o fylogenetickém původu, době vzniku a šíření subsaharských haploskupin mtDNA vznikla na základě výzkumu asi 2700 vzorků nejen z afrických nemocnic a transfuzních center, ale i od Afroameričanů žijících v Jižní a Severní Americe. Databáze evropských sekvencí mtDNA, která počet afrických vzorků několikanásobně přesahuje, jasně naznačuje, že velký africký kontinent skrývá ještě mnohá překvapení. 8)
Současná představa o fylogenetickém původu, době vzniku a šíření subsaharských haploskupin mtDNA vznikla na základě výzkumu asi 2700 vzorků nejen z afrických nemocnic a transfuzních center, ale i od Afroameričanů žijících v Jižní a Severní Americe. Databáze evropských sekvencí mtDNA, která počet afrických vzorků několikanásobně přesahuje, jasně naznačuje, že velký africký kontinent skrývá ještě mnohá překvapení. 8)
Výzkum mtDNA v Čadské pánvi
Výzkum rozmanitosti (diverzity) mtDNA, který v rámci grantového projektu uskutečnili na území Čadské pánve v letech 2002–2004 badatelé Archeologického ústavu AV ČR v Praze, Ústavu hematologie a krevní transfuze a Fakulty humanitních studií Univerzity Karlovy v Praze, ukázal, že úspěchu lze dosáhnout zejména v odlehlých oblastech. Zde totiž žijí lidé, kteří větší města s lékařskou péčí obvykle nenavštěvují a o jejichž mtDNA bývají analýzy populačních genetiků ochuzeny. V africkém sahelu, oblasti související s pravěkými břehy Čadského jezera, jsme získali vzorky od několika zemědělských a pasteveckých populací. Zvláštní pozornost jsme věnovali společnostem čadské větve afroasijských jazyků, jejichž mtDNA nebyla do té doby ještě publikována. Získali jsme vzorky od Budumů z ostrovů v Čadském jezeře (obrázek 1 a obrázek 8), Kotoků z povodí Šari, Masů z povodí Logon (obrázek 6) a Mafů z pohoří Mandara. Kromě čadských skupin byli vzorkováni i zástupci nilosaharské (Kanuri a Kanembu) a nigerokonžské (Fali a Fulbové) jazykové rodiny (obrázek 7). Vzorky jsme získali také od kočovných a polokočovných arabských kmenů. Naše databáze dnes obsahuje 448 vzorků od vzájemně nepříbuzných jedinců.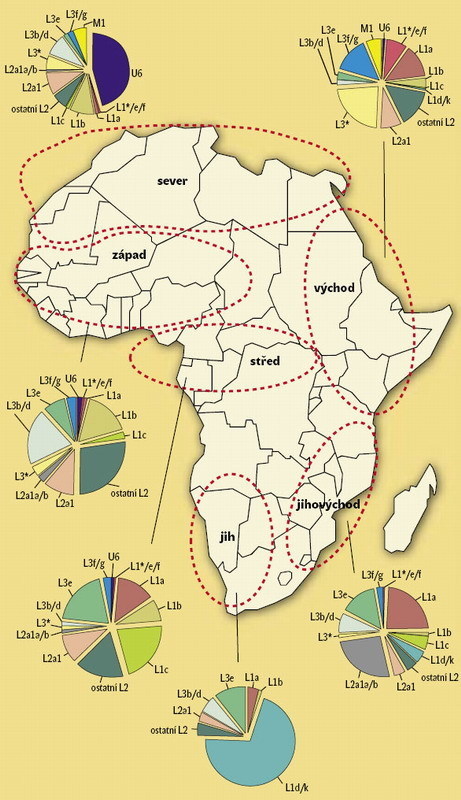 V populačním vzorku z Čadské pánve jsme našli nejen publikované a známé subsaharské haploskupiny, ale i sporadicky se vyskytující haploskupiny euroasijské (M, M1, R, pre-HV, U3, U4, U5 a V), které měly především pastevecké populace (Arabů a Fulbů). Zaujala nás však haploskupina L3f, která se ve fylogenetické síti dále rozpadala do dosud neznámé podoby. Tuto skupinu jsme nazvali L3f2 (viz obrázek 9). Z antropologického hlediska je velmi zajímavé, že se L3f2 vyskytuje téměř výhradně u čadských skupin, přestože jsou v našem vzorku zastoupeny pouze z 38 %. Znamená to, že tato haploskupina vypovídá nejspíše o populacích hovořících čadskými jazyky.
V populačním vzorku z Čadské pánve jsme našli nejen publikované a známé subsaharské haploskupiny, ale i sporadicky se vyskytující haploskupiny euroasijské (M, M1, R, pre-HV, U3, U4, U5 a V), které měly především pastevecké populace (Arabů a Fulbů). Zaujala nás však haploskupina L3f, která se ve fylogenetické síti dále rozpadala do dosud neznámé podoby. Tuto skupinu jsme nazvali L3f2 (viz obrázek 9). Z antropologického hlediska je velmi zajímavé, že se L3f2 vyskytuje téměř výhradně u čadských skupin, přestože jsou v našem vzorku zastoupeny pouze z 38 %. Znamená to, že tato haploskupina vypovídá nejspíše o populacích hovořících čadskými jazyky.
Molekulární datování L3f2 včetně další dosud nepublikované haploskupiny L3e5 ukazuje na dobu, kdy ve zkoumané oblasti končila sušší klimatická fáze, tzv. kanemien. Paleoklimatologické a archeologické výzkumy vypovídají, že v následujícím holocenním období nastoupilo výrazně vlhčí klima a vzniklo ohromné Megačadské jezero, k němuž se jistě stahovaly populace hledající obživu. Nejspíš to byli pastevci, kteří si svůj jídelníček zpestřovali lovem zvěře a ryb. Některá lingvistická srovnání čadských a kušitských jazyků naznačují, že protočadské populace sem mohly přicházet z nilského údolí, z oblasti ležící v úrovni 3.–4. kataraktu. Pro kolonizaci východních oblastí Čadské pánve musely využívat vysychající řeky se stojatou zbytkovou vodou.
Archeologické výzkumy v západním Súdánu dokládají putování protočadských pastevců údolím Howar. Možná že k populační expanzi, která se dnes projevuje hvězdicovitým uspořádáním haploskupin L3f2 a L3e5, mohlo dojít právě zde. Protože tyto skupiny nebyly dosud ve východní Africe zjištěny, dalo by se soudit, že se protočadští pastevci krátce po své diverzifikaci odebrali k Čadskému jezeru, kde je stále sušší a sušší klima oddělilo od příbuzných východoafrických (kušitských) skupin.
Naše výzkumy ukázaly, jak ještě málo známe mtDNA afrických populací. Je velmi pravděpodobné, že geneticky neprozkoumaná území mezi Čadským jezerem a nilským údolím poskytnou nové sekvence mtDNA, které poodhalí roušku historie tohoto krásného, ale dosud skrytého koutu planety. Kolik měla naše pramatka Eva afrických dcer, zatím přesně nevíme – ale jejich současný počet se v brzké době nejspíš ještě trochu zvýší.
Výzkum v Čadské pánvi se uskutečnil díky podpoře Grantové agentury České republiky (404/03/0318).
Poznámky
 V brzké době vyjde v nakladatelství Academia autorova kniha Lidé od Čadského jezera. Zabývá se otázkou, jakou úlohu hrálo Čadské jezero při vzniku etnických skupin sahelosúdánského pásma Afriky ležícího mezi Saharou a tropickými deštnými pralesy. Autor využívá jak společenskovědní, tak přírodovědné prameny, které doplňuje vlastními poznatky z antropologických expedic realizovaných během posledních pěti let. V šesti tematicky různých kapitolách sleduje historické záznamy arabských a evropských cestovatelů, vývoj přírodního prostředí, osudy tamních civilizací a kultur, současnou etnickou skladbu, specifika vybraných společenských skupin a v závěru pojednává o biologických aspektech místních obyvatel. Autor, který měl možnost opřít svůj výklad osídlení Čadské pánve o vlastní molekulárněgenetické analýzy, vyvozuje závěry o migracích a původu jednotlivých populačních skupin se zvláštním zřetelem na čadské zemědělce a fulbské pastevce. Kniha ukazuje, že neutěšené prostředí stále pulzujícího Čadského jezera vytváří jakýsi mezník řady kulturních vlivů i populačních celků přicházejících sem ze západní i východní části sahelosúdánského prostoru; podmiňuje ale zároveň i vznik specifických rysů, které rozšiřují naši mlhavou představu o lidech této opomíjené části dynamicky se měnícího světa.
V brzké době vyjde v nakladatelství Academia autorova kniha Lidé od Čadského jezera. Zabývá se otázkou, jakou úlohu hrálo Čadské jezero při vzniku etnických skupin sahelosúdánského pásma Afriky ležícího mezi Saharou a tropickými deštnými pralesy. Autor využívá jak společenskovědní, tak přírodovědné prameny, které doplňuje vlastními poznatky z antropologických expedic realizovaných během posledních pěti let. V šesti tematicky různých kapitolách sleduje historické záznamy arabských a evropských cestovatelů, vývoj přírodního prostředí, osudy tamních civilizací a kultur, současnou etnickou skladbu, specifika vybraných společenských skupin a v závěru pojednává o biologických aspektech místních obyvatel. Autor, který měl možnost opřít svůj výklad osídlení Čadské pánve o vlastní molekulárněgenetické analýzy, vyvozuje závěry o migracích a původu jednotlivých populačních skupin se zvláštním zřetelem na čadské zemědělce a fulbské pastevce. Kniha ukazuje, že neutěšené prostředí stále pulzujícího Čadského jezera vytváří jakýsi mezník řady kulturních vlivů i populačních celků přicházejících sem ze západní i východní části sahelosúdánského prostoru; podmiňuje ale zároveň i vznik specifických rysů, které rozšiřují naši mlhavou představu o lidech této opomíjené části dynamicky se měnícího světa.
(250 stran, 97 čb fotografií v textu, 32 stran barevné přílohy, vázané.)
Ke stažení
 článek ve formátu pdf [2,44 MB]
článek ve formátu pdf [2,44 MB]
O autorovi
Viktor Černý

Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
