










Ernst Mayr a teorie přerušovaných rovnováh
 O jedno z největších překvapení v evoluční biologii 20. století se postarali paleontologové Stephen Jay Gould a Niels Eldredge, kteří v šedesátých letech publikovali teorii přerušovaných rovnováh (punktuační teorii evoluce). Podle ní se druhy nevyvíjejí tak, jak si představoval Darwin, tj. postupným rovnoměrným hromaděním drobných změn, gradualisticky, ale ve skocích časově vázaných na okamžiky vzniku druhů, punktualisticky (obrázek). Paleontologický záznam prý ukazuje, že se většina druhů vyvíjí, tj. mění své vlastnosti, pouze v krátkém období po svém vzniku (tisíce až desetitisíce let), zatímco po zbytek své existence (miliony let) zůstávají víceméně neměnné. Teorie přerušovaných rovnováh byla nejdřív – jako každý zásadní objev – většinou paleontologů a evolučních biologů odmítnuta, postupem času však přibývala data nasvědčující tomu, že by uvedený model mohl být správný. A ačkoli se to běžně neví, jedním z duchovních otců teorie přerušovaných rovnováh byl i Ernst Mayr.
O jedno z největších překvapení v evoluční biologii 20. století se postarali paleontologové Stephen Jay Gould a Niels Eldredge, kteří v šedesátých letech publikovali teorii přerušovaných rovnováh (punktuační teorii evoluce). Podle ní se druhy nevyvíjejí tak, jak si představoval Darwin, tj. postupným rovnoměrným hromaděním drobných změn, gradualisticky, ale ve skocích časově vázaných na okamžiky vzniku druhů, punktualisticky (obrázek). Paleontologický záznam prý ukazuje, že se většina druhů vyvíjí, tj. mění své vlastnosti, pouze v krátkém období po svém vzniku (tisíce až desetitisíce let), zatímco po zbytek své existence (miliony let) zůstávají víceméně neměnné. Teorie přerušovaných rovnováh byla nejdřív – jako každý zásadní objev – většinou paleontologů a evolučních biologů odmítnuta, postupem času však přibývala data nasvědčující tomu, že by uvedený model mohl být správný. A ačkoli se to běžně neví, jedním z duchovních otců teorie přerušovaných rovnováh byl i Ernst Mayr.
Čím přerušovaný průběh evoluce vysvětlovali?
Eldredge a Gould zpočátku navrhli dvě možnosti. První z nich (podle mého názoru správnou) však opustili a přiklonili se k druhé, méně radikální. I mladoturci časem zestárnou…- Podle první hypotézy je za evoluční schopnost proměny nově vznikajícího druhu odpovědná genetická revoluce, jak ji popsal v jednom ze svých zásadních děl Živočišné druhy a evoluce právě Ernst Mayr. 1) Nepočetná populace, která se při peripatrické speciaci odštěpí od populace velké, si s sebou odnese jen malou část celkového genetického polymorfizmu. To může vyvolat v genetické skladbě nové populace další prudké změny, jež mohou být provázeny odpovídajícími změnami ve vzhledu příslušníků nového druhu. Zatímco ve velké populaci byla měřítkem evoluční úspěšnosti alely (varianty genu) její schopnost uspokojivě fungovat ve společnosti co největšího počtu alel vlastního genu i alel cizích genů, v odštěpené populaci, v níž zůstala pouze malá část původního genetického polymorfizmu, nabývá na významu míra, v jaké daná alela ovlivňuje biologickou zdatnost svého nositele. Mayr pochopitelně ve čtyřicátých letech minulého století nemohl vědět o teorii evolučně stabilních strategií, z níž dnes vyplývá, že geneticky polymorfní populace pohlavně se rozmnožujícího druhu může snadno „evolučně zamrznout“ a přestat odpovídat na selekční tlak. Intuice a dobrý pozorovací talent mu zřejmě napověděly, že genetické revoluce, díky nimž při některých typech vzniku druhů začne evolučně zamrzlá populace na selekční tlaky znovu odpovídat, by mohly hrát v procesu biologické evoluce zásadní roli.
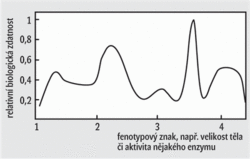
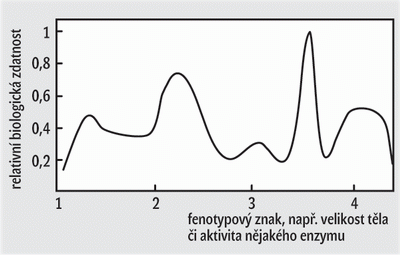
- Podle druhé hypotézy lze průběh evoluce ve skocích vysvětlit výrazným zmenšením populace, které provází každý vznik druhu vyvolaný odštěpením malé části populace z populace velké. Již Sewall Wright, jeden z otců zakladatelů neodarwinizmu, zdůrazňoval, že přechodné zmenšení populace, které je nutně provázeno výrazným snížením účinnosti jakéhokoli výběru, může paradoxně adaptivní evoluci zefektivnit. Pouze za situace, kdy přirozený výběr na čas sníží svou účinnost, mohou přežít a zanechat potomky mutovaní jedinci s přechodnými vlastnostmi, tj. takoví, kteří již nejsou přizpůsobeni využívání ekologické niky původního druhu a zároveň ještě nejsou dobře přizpůsobeni využívání niky druhu nového. Wright to popisoval pomocí modelu adaptivní krajiny (obrázek): Ve velké populaci nemají příliš šanci na přežití mutovaní jedinci, kteří se nacházejí na svazích kopců, nebo dokonce hluboko v údolích. Bez přechodného sestupu do údolí však není možné obsadit v adaptivní krajině nové vrcholy. Wrightův model posunující se rovnováhy, jímž se vysvětluje úloha přechodného zmenšení populace, je poněkud složitější, než jak zde byl představen. V důsledku toho a patrně i vinou náročného matematického aparátu, který Wright ve svých pracích obvykle užíval (kdo by se těmi integrály prokousával, pojďme radši měřit šupiny cichlidám), upadal tento model postupně mezi evolučními biology v nemilost. Možná i proto se na něj Eldredge s Gouldem příliš neodvolávali a spíše zdůrazňovali některé další výhody méně početných populací:
 Malých populací vzniká při hranicích areálů mnoho a každá z nich představuje samostatný evoluční experiment, což podstatně zvyšuje šanci, že nový druh vznikne právě z malé populace. V malých populacích probíhají veškeré evoluční změny rychleji než ve velké. Malé populace se vyskytují při hranicích areálu velké populace, to znamená v podmínkách z hlediska druhu extrémních. Příslušníci malých populací jsou také vystaveni odpovídajícím extrémním selekčním tlakům. Populace nově vznikajících druhů se vyskytují na omezeném území. Na něm patrně panují ve srovnání s hlavním areálem výskytu mateřského druhu všude stejné nebo alespoň podobné podmínky, a tak jsou všude stejné i selekční tlaky působící na příslušníky vznikajícího druhu. Izolovanost od areálu mateřské populace zároveň zajišťuje, že nikdo nepřináší do genofondu cizí alely, které by svou přítomností zhoršovaly šanci populace přizpůsobit se místním podmínkám. Díky současnému vlivu všech těchto faktorů není vlastně divu, že úspěšný evoluční experiment – vznik nového druhu, podstatně odlišného od druhu původního – proběhne téměř pokaždé právě v některé málo početné populaci v blízkosti hlavního areálu mateřského druhu (a nikoli v početné populaci uvnitř hlavního areálu). To, co nakonec vidíme v paleontologickém záznamu jako bleskové vystřídání starého druhu novým, není ve skutečnosti evoluční děj vzniku nového druhu v daném místě, ale děj ekologický – vytěsnění starého druhu druhem novým, který vznikl někdy dříve a někde jinde a v daný okamžik pouze expandoval na úkor areálu původního druhu.
Malých populací vzniká při hranicích areálů mnoho a každá z nich představuje samostatný evoluční experiment, což podstatně zvyšuje šanci, že nový druh vznikne právě z malé populace. V malých populacích probíhají veškeré evoluční změny rychleji než ve velké. Malé populace se vyskytují při hranicích areálu velké populace, to znamená v podmínkách z hlediska druhu extrémních. Příslušníci malých populací jsou také vystaveni odpovídajícím extrémním selekčním tlakům. Populace nově vznikajících druhů se vyskytují na omezeném území. Na něm patrně panují ve srovnání s hlavním areálem výskytu mateřského druhu všude stejné nebo alespoň podobné podmínky, a tak jsou všude stejné i selekční tlaky působící na příslušníky vznikajícího druhu. Izolovanost od areálu mateřské populace zároveň zajišťuje, že nikdo nepřináší do genofondu cizí alely, které by svou přítomností zhoršovaly šanci populace přizpůsobit se místním podmínkám. Díky současnému vlivu všech těchto faktorů není vlastně divu, že úspěšný evoluční experiment – vznik nového druhu, podstatně odlišného od druhu původního – proběhne téměř pokaždé právě v některé málo početné populaci v blízkosti hlavního areálu mateřského druhu (a nikoli v početné populaci uvnitř hlavního areálu). To, co nakonec vidíme v paleontologickém záznamu jako bleskové vystřídání starého druhu novým, není ve skutečnosti evoluční děj vzniku nového druhu v daném místě, ale děj ekologický – vytěsnění starého druhu druhem novým, který vznikl někdy dříve a někde jinde a v daný okamžik pouze expandoval na úkor areálu původního druhu.
Role genetické revoluce
Domnívám se, že svůj nápad s klíčovou rolí genetické revoluce zavrhli Eldredge s Gouldem předčasně. 2) Myslím, že šlo o exemplární ukázku metodicky správného, věcně však chybného použití Occamovy břitvy. 3) Occamovo doporučení, že se entity nemají zbytečně zmnožovat, se dnes většinou tlumočí jako velmi rozumný požadavek, že ze dvou modelů, které podstatu jevu vysvětlují stejně dobře, musí dát vědec přednost tomu jednoduššímu. Ne že by si vědci naivně mysleli, že příroda je jednoduchá. Modely (hypotézy) jsou však určeny v první řadě k tomu, abychom se je pokoušeli vyvracet. A jednoduché modely lze vyvrátit snáze než modely složitější. 4) Eldredge s Gouldem se patrně domnívali, že pro vysvětlování skoků v paleontologickém záznamu vystačí s všeobecně přijímaným jevem vzniku druhů z drobných populací, a že tedy do svého modelu nemusí zahrnovat poněkud exotický jev genetických revolucí, o kterém ostatně většina evolučních biologů neslyšela a ti zbývající v něj většinou nevěří. Z metodologického i taktického hlediska šlo určitě o správné rozhodnutí. Jednoduchý model opírající se výhradně o působení obecně známých a všeobecně uznávaných procesů má samozřejmě mnohem větší šanci na pozitivní přijetí než složitější model, předpokládající navíc působení procesu, jehož existence je v lepším případě předmětem diskuse, v horším případě objektem kolektivního zapomnění. Z věcného hlediska však šlo, alespoň podle mého názoru, o rozhodnutí chybné. Jednoduchý slovní model může sice zdánlivě vysvětlit skoky v paleontologickém záznamu, řadu jiných jevů však nevysvětlí.Evoluce u sexuálně se rozmnožujících druhů
Jednoduchý model například nemůže vysvětlit samotnou existenci adaptivní evoluce u sexuálně se rozmnožujících druhů, u kterých by podle teoretických modelů k evoluci vůbec nemělo docházet. Nebo takový detail, který se čím dál jasněji rýsuje při studiu výskytu rovnoměrné a přerušované evoluce různých skupin organizmů: Opakovaně bylo zjištěno, že u mořských jednobuněčných organizmů se vyskytuje spíše rovnoměrná evoluce, u mořských mnohobuněčných organizmů probíhá spíše ve skocích. V nepočetných populacích oddělených od populace velké by přitom druhy měly vznikat jak u jednobuněčných, tak u mnohobuněčných. U jednobuněčných organizmů se ovšem také v mnohem větší míře setkáváme s nepohlavním rozmnožováním. Je proto klidně možné, že častější výskyt rovnoměrné evoluce u jednobuněčných souvisí s jejich častou schopností nepohlavního množení, tj. se schopností evolučně nezamrzat, a tedy podléhat klasické kontinuální darwinistické evoluci.Dokud nebude jednoznačně prokázáno, že v malých izolovaných populacích skutečně evoluce probíhá rychleji (navzdory nízké účinnosti selekce a malému počtu nových mutací, které zde mohou vzniknout), a dokud pro existenci adaptivní evoluce u sexuálně se rozmnožujících druhů a pro postupný rovnoměrný průběh evoluce u jednobuněčných organizmů nebudou navržena jiná vysvětlení, považoval bych odmítnutí účasti genetických revolucí při vysvětlování přerušovaného charakteru evoluce za přinejmenším, hmm, poněkud předčasné. To jsem nakonec smířlivý, co?
Poznámky
ERNST MAYR
/*5. 7. 1904 Kempten v Německu, †3. 2. 2005 Bedford v Massachusetts/ Z jihoněmeckého Kemptenu se s rodiči přestěhoval do Würzburgu a později do Mnichova, kam byl jeho otec přeložen jako státní návladní. Po otcově smrti se v předposledním roce první světové války (1917) přestěhoval s matkou a dvěma bratry do Drážďan. R. 1923 začal studovat medicínu na Lékařské fakultě Univerzity v Greifswaldu a po složení státních zkoušek r. 1925 studoval zoologii na Univerzitě v Berlíně. Ke změně oboru jej přemluvil jeden z nejvlivnějších ornitologů 20. století Erwin Stresemann (1889–1972) tím, že mu slíbil účast na výzkumné expedici. Po promoci (r. 1926) se Mayr stal asistentem v Zoologickém muzeu v Berlíně, kde působil i Stresemann. O dva roky později sám podnikl výpravu do tehdejší nizozemské kolonie na Nové Guinei, jež se protáhla tím, že se přidal k expedici týmu Amerického přírodovědeckého muzea z New Yorku na Šalamounovy ostrovy (Whitney Expedition). Právě v tomto muzeu pak r. 1932 nastoupil jako pomocný kurátor, aby uspořádal materiál ze zmíněné expedice. Potom však muzeum odkoupilo proslulou ptačí sbírku Rothschildů (měla 280 000 exemplářů) a Mayrovi bylo uloženo zpracovat ji a začlenit do sbírek. Tento jeho úkol se protáhl až do druhé světové války. R. 1944 se stal řádným kurátorem muzea. Po válce zůstal ve Spojených státech a koncem r. 1950 získal americké občanství. R. 1935 se oženil s Margarete Simonovou (1912–1990), s níž měl dcery Christu a Susanne.Zemřel po krátké nemoci v Bedfordu. Loni 5. července stačil ještě oslavit své sté narozeniny. Byl jedním z nejvýznamnějších evolučních biologů 20. století, předním ornitologem a systematikem, bojovným obhájcem nové filozofie biologie a jedním z nejvlivnějších historiků biologie.
Z obsáhlého Mayrova díla vybíráme:
Systematics and the Origin of Species, Columbia University Press, New York 1942
Animal Species and Evolution, Harvard University Press, Cambridge 1963
The Growth of Biological Thought. Diversity, Evolution, and Inheritance, The Belknap Press and Harvard University Press, Cambridge and London 1982
This is Biology: The Science of the Living World, Harvard University Press, Cambridge 1997
(spolu s Jaredem Diamondem) The Birds of Northern Melanesia, Oxford University Press, Oxford and New York 2001
adaptace – změny vzniklé tím, že se organizmus evolučně přizpůsobil podmínkám prostředí
adaptivní evoluce – organizmy se postupně, řadou dílčích kroků přizpůsobují prostředí působením selekce
evolučně stabilní strategie – když nositelé vlastnosti v populaci převládnou, nemohou být selekčně vytlačeni nositeli jiné, menšinové strategie
genetická revoluce – v Mayrově pojetí: odštěpená malá populace si z velké populace odnese jen malou část celkového genetického polymorfizmu. To může vyvolat v genetické skladbě nové populace prudké změny, jež mohou být provázeny odpovídajícími změnami ve vzhledu příslušníků nového druhu.
genetický polymorfizmus – genetická různorodost v populaci. U nepohlavně se rozmnožujících organizmů se dědí genetická výbava (genotyp) z generace na generaci a nositelé nevýhodných genotypů jsou v procesu přirozeného výběru z populace odstraňováni i s genotypem. U pohlavně se rozmnožujících organizmů vznikají nové genotypy jako kombinace genů od obou rodičů, a tím se v každé generaci obnovují i genotypy nevýhodné. Ty se však za změněných vnějších podmínek mohou ukázat jako výhodné. Pohlavně se rozmnožující druh si tedy dokáže v skrytu uchovat určité množství polymorfizmu, které mu umožňuje rychleji se evolučně přizpůsobit měnícím se přírodním podmínkám.
syntetická teorie evoluce – neodarwinizmus. Když byly znovuobjeveny Mendelovy zákony a vznikla genetika, evolucionisté se pokusili Mendela s Darwinem skloubit. Genostředné pojetí evoluce W. D. Hamiltona, G. Williamse a R. Dawkinse stejně jako eklektické pojetí evoluce J. S. Goulda z poslední třetiny 20. století již rámec neodarwinizmu překračují.
teorie evolučně stabilních strategií – výsledek práce J. Maynarda Smitha a G. Priceho. Teorie ukazuje, že v evoluci nakonec nepřevládnou alely poskytující svým nositelům nejvyšší biologickou zdatnost, ale alely podmiňující evolučně stabilní strategii.
teorie přerušovaných rovnováh – punktuační teorie. N. Eldredge a J. S. Gould předpokládají, že druhy mění své vlastnosti pouze v době bezprostředně po svém vzniku a následně zůstávají po mnohem delší období (až do svého vymření) neměnné.
teorie sobeckého genu – teorie mezialelické selekce. Poukazuje na skutečnost, že u pohlavně se rozmnožujících druhů se genotyp, a tedy ani biologická zdatnost nedědí, tudíž tyto organizmy nemohou být výtvorem darwinovské evoluce. Subjektem evoluce jsou jednotlivé alely stejného genu, které mezi sebou soutěží o to, v kolika kopiích se dokážou přenést do dalších generací.
teorie zamrzlé plasticity – geneticky polymorfní populace pohlavně se rozmnožujícího druhu může velmi snadno „evolučně zamrznout“ a přestat odpovídat na selekční tlak. Teorie vychází ze stejných argumentů jako teorie sobeckého genu, navíc však uvažuje i existenci mezigenových (epistatických) interakcí a bere v úvahu i závěry teorie evolučně stabilních strategií.
Ke stažení
 článek ve formátu pdf [275,21 kB]
článek ve formátu pdf [275,21 kB]
O autorovi
Jaroslav Flegr

Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
