










Spor o mitochondrie
Klasifikace nejjednodušších eukaryotických organizmů, protistů, byla založena až do poloviny osmdesátých let převážně na morfologii, zejména na srovnávacím studiu stavby buněk a jejich složek. Stejně na tom bylo i odhalování jejich fylogenetických vztahů a spekulace o stáří jednotlivých skupin, posuzovaném podle stupně složitosti buněčné organizace. Přitom bylo nutné překonávat mnoho těžkostí, a tak se určování fylogenetické původnosti a pokročilosti stále podobalo chůzi v mlze, z níž jen tu a tam vyvstávaly ostrůvky určitých tvarů.
Fascinující možnosti nabídlo srovnávání sekvencí malých podjednotek ribozomální RNA (SSU rRNA), těchto „jedinečných fylogenetických hodin“ (C. R. Woese), které umožňovalo zjistit vzájemné fylogenetické postavení jednotlivých skupin protistů, a bylo proto uvítáno s obrovským zájmem (viz též Z. Storchová, Vesmír 76, 612, 1997/11). Překvapivá zjištění na sebe nedala dlouho čekat. Práce týmů M. L. Sogina a C. R. Vossbrincka v USA ukázaly, že podle sekvencí podjednotek rRNA se jako první z vývojové linie eukaryontů odvětvily tři skupiny protistů: bičíkovci metamonády zastoupení známým rodem Giardia, nitrobuněční paraziti mikrosporidie (první byla osekvencována Vairimorpha) a bičíkovci skupiny Parabasalia (Trichomonas).
Tři skupiny protistů
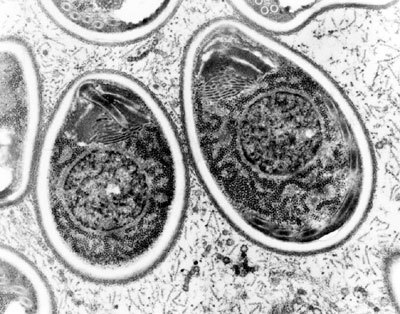
Stručně tyto organizmy představme: Giardia intestinalis (obrázek a obrázek), běžný střevní parazit člověka, který může ve střevě vyvolat zánět, patří k nejvyvinutějším zástupcům metamonád z řádu Diplomonadida, kteří představují dvojice natrvalo splynuvších jedinců s celkem osmi bičíky. Mikrosporidie (obrázek a obrázek) jsou známy jako cizopasníci téměř všech skupin živočichů, zejména jako patogeni u hmyzu a ryb.{kind=link}
{kind=link}
{kind=link}
{kind=link}
 V poslední době na sebe soustředili pozornost jako paraziti člověka u pacientů trpících aidsem. Mají spory s jedinečným zařízením, vystřelovacím aparátem, jímž vstřikují infekční zárodek, sporoplazmu, jakoby injekční jehlou pod velkým tlakem do nového hostitele (podrobněji viz Vesmír 72, 578, 1993/10). Trichomonas (obrázek a obrázek), nejznámější rod parabazálií, zahrnuje parazitické bičíkovce převážně ze zažívacího traktu různých živočichů včetně člověka (např. T. vaginalis v urogenitálním traktu), má několik funkčně specializovaných bičíků, undulující membránu (lat. unda – vlna, undare – vlnit) a složitý mikrotubulární cytoskelet.
V poslední době na sebe soustředili pozornost jako paraziti člověka u pacientů trpících aidsem. Mají spory s jedinečným zařízením, vystřelovacím aparátem, jímž vstřikují infekční zárodek, sporoplazmu, jakoby injekční jehlou pod velkým tlakem do nového hostitele (podrobněji viz Vesmír 72, 578, 1993/10). Trichomonas (obrázek a obrázek), nejznámější rod parabazálií, zahrnuje parazitické bičíkovce převážně ze zažívacího traktu různých živočichů včetně člověka (např. T. vaginalis v urogenitálním traktu), má několik funkčně specializovaných bičíků, undulující membránu (lat. unda – vlna, undare – vlnit) a složitý mikrotubulární cytoskelet.
{kind=link}
{kind=link}
Všechny tři skupiny postrádají mitochondrie – organely obsahující zejména oxidační enzymy a plnící roli jakési buněčné „elektrárny“. To se pěkně shodovalo s představou velmi dávných forem, pocházejících ještě z doby před symbiotickým vznikem mitochondrií. Možnost druhotné ztráty mitochondrií se odsouvala do pozadí. Trochu ale udivovalo, že všechny tři „rané“ skupiny jsou už vysoce uzpůsobené k parazitizmu (mikrosporidie dokonce k výhradně nitrobuněčnému) a že žijí v hostitelích, kteří vznikli nesrovnatelně později než jejich paraziti. To se dalo vysvětlit tak, že dávní amitochondriální a ještě volně žijící eukaryonti se zachraňovali před stoupající hladinou kyslíku v prostředí tím, že se uchýlili do bezkyslíkatých prostředí uvnitř hostitele. Adaptace jejich potomků k parazitizmu by pak vysvětlila jejich nyní velice pokročilou stavbu těla, jako je složitý cytoskelet metamonád i parabazálií nebo velmi důmyslné „injekční“ zařízení mikrosporidií.
Tyto tři skupiny, údajné potomky premitochondriálních eukaryont, zařadil už r. 1983 T. Cavalier-Smith jako kmeny do říše Archezoa, kterou nově ustavil. K nim přidal ještě problematicky různorodou skupinu Archamoebae (ve složeninách s první částí pocházející z ř. archaios – starý se mu zalíbilo).  Do ní zařadil rody měňavkovitých organizmů, které nemají mitochondrie, jako je velká, mnohojaderná, volně žijící měňavka Pelomyxa nebo střevní parazit zvířat i člověka Entamoeba. Některé další charakteristiky také svědčí o tom, že uvedené tři skupiny se oddělily od hlavní linie vedoucí k vyšším eukaryontům už v rané fázi eukaryotické evoluce (z toho mikrosporidie jako první). Mikrosporidie mají nadto některé vlastnosti, jimiž se podobají prokaryontům – ribozomy s charakteristikou 70 S (nejsou však samy – mj. je mají i Parabasalia – např. Trichomonas) a nejmenší známé genomy, většinou pod 20 megabází, v některých případech dokonce menší než u bakterií. Mikrosporidie druhu Encephalitozoon cuniculi, cizopasník savců, má genom o méně než 2,9 megabáze.
Do ní zařadil rody měňavkovitých organizmů, které nemají mitochondrie, jako je velká, mnohojaderná, volně žijící měňavka Pelomyxa nebo střevní parazit zvířat i člověka Entamoeba. Některé další charakteristiky také svědčí o tom, že uvedené tři skupiny se oddělily od hlavní linie vedoucí k vyšším eukaryontům už v rané fázi eukaryotické evoluce (z toho mikrosporidie jako první). Mikrosporidie mají nadto některé vlastnosti, jimiž se podobají prokaryontům – ribozomy s charakteristikou 70 S (nejsou však samy – mj. je mají i Parabasalia – např. Trichomonas) a nejmenší známé genomy, většinou pod 20 megabází, v některých případech dokonce menší než u bakterií. Mikrosporidie druhu Encephalitozoon cuniculi, cizopasník savců, má genom o méně než 2,9 megabáze.
Kmen Parabasalia (Trichomonas) však v říši Archezoa dlouho nepobyl. U Archezoí sice chybějí typické mitochondrie, ale zato mají organely s jednoduchou či dvojitou membránou (známé už od sedmdesátých let a zřejmě postrádající vlastní genom), hydrogenozomy. Jsou vybaveny oxidačními enzymy spojenými s produkcí ATP, nikoliv sice oxidativní fosforylací, ale konverzí pyruvátu či malátu na acetát, CO2 a plynný vodík (odtud jejich jméno). Bylo předmětem dlouhých diskusí, zda jsou to přeměněné mitochondrie, nebo zda mají původ v endosymbiontech či snad v diferenciaci endomembránových systémů. Nejsou přítomny jen u parabazálií, ale i u některých nálevníků a měňavkovitých a houbových organizmů. Nevystupují u nich nikdy spolu s mitochondriemi – např. ti nálevníci, kteří mají hydrogenozomy, nemají mitochondrie. To mluvilo spíš pro původ z přeměněných mitochondrií, a proto je Cavalier-Smith r. 1987 přeřadil do říše protozoí.
Rodokmen eukaryotických organizmů
S postupem času byly poznány sekvence podjednotek ribozomální rRNA u několika set eukaryotických organizmů, takže bylo možno sestavit podle nich jakýsi jejich rodokmen. Tento hypotetický strom je tvořen třemi doménami: základní, kam patří ony tři amitochondriální skupiny a z níž jako z kořenů vyrůstá „kmen“ tvořený skupinou bičíkovců Euglenozoa (kam patří známé volně žijící krásnoočko, Euglena gracilis, ale také cizopasné trypanozomy, včetně původců spavé nemoci) i nejrůznější měňavkovité organizmy. Třetí je velmi široká „terminální koruna“, která zahrnuje ohromnou škálu protistů a také tři říše mnohobuněčných – rostliny, živočichy a houby. Tomu odpovídaly názory na to, v jaké posloupnosti se u protistů objevovaly buněčné organely. Mikrosporidie v základní doméně mají jen endoplazmatické retikulum, archaméby a metamonády o něco výše nad ní mají už bičíky a bazální tělíska, a u parabazálií se už objevuje Golgiho aparát. Skutečné mitochondrie a chloroplasty jako výsledek endosymbiózy s prokaryotickými organizmy se objevují teprve u protistů na úrovni „kmenu“.
Ještě nežli uvedené představy o skupině archezoí jako prapůvodních eukaryontech vešly v širší známost a začaly pronikat do učebnic, ukázalo srovnání sekvencí podjednotky SSU rRNA, že zástupci archaméb, Entamoeba a Pelomyxa, se odvětvují od hlavní linie až spolu s eukaryonty vybavenými mitochondriemi. Asi tedy mitochondrie mohly mít a pak je ztratit podobně jako pravděpodobně parabazália. Ještě průkaznější byl pozdější nález dvou mitochondriálních genů kódujících bílkoviny v genomu entamoeby (pyridin-nukleotid transhydrogenázy a chaperoninu cpn60), kam se dostaly přenosem z genomu původního symbionta. To byl prvý test pravdy pro říši Archezoa, ve kterém její původnost neobstála.
Pozůstatky mitochondrií byly pak nalezeny i u metamonád. Je nutné říci, že vše, co přispívá k poznání fylogeneze této skupiny, bylo objeveno u rodu Giardia. V loňském roce podaný průkaz chaperoninového genu 60, podobného mitochondriálním genům, silně svědčí také o někdejší existenci endosymbiontů, patrně předků mitochondrií, v buňce metamonád.
Mají mikrosporidie společné předky s houbami?
Zvlášť silně je novými nálezy ovlivněno postavení mikrosporidií. Konečně je možno mít za prokázané, že struktury dlouho pokládané za „primitivní“ Golgiho aparát jím skutečně jsou, i s diferenciací podobnou té u vyšších eukaryontů. To není nic zvláštního vzhledem k překvapivým zjištěním, která naznačují, že se mikrosporidie mohly vyvinout z „terminální koruny“ eukaryontů a patrně sdílejí společné předky s houbami. Geny bílkovin elongačního faktoru mají totiž vloženou sekvenci, která je společná mikrosporidiím, živočichům a houbám a některé enzymy (dihydrofolátreduktáza a thymidylátsyntáza) mají u těchto tří skupin strukturu odlišnou od ostatních eukaryontů. Ještě důležitější je nález mitochondriálních chaperoninových genů cpn70 v genomech mikrosporidií rodů Nosema a Vairimorpha. Pro příbuznost mikrosporidií s houbami zprvu mluvily jen podobnosti neobvyklého průběhu meiotického cyklu mikrosporidií a některých hub, ale nyní je tato domněnka posílena fylogenezí bílkovin α a β tubulinu, jimiž se mikrosporidie řadí do fylogenetické radiace hub. Další podporou je, že ve fylogenetických rodokmenech se sekvence genů cpn70 větví, i když slabě, spolu se sekvencemi mitochondriálních genů hub rodu Boveria. Mikrosporidie tedy s velkou pravděpodobností nejsou potomci prastarých nebo primitivních eukaryontů, ve velké míře pozměněných adaptacemi k parazitizmu, ale naopak vzešly z organizmů značně pokročilých.
Pro příbuznost mikrosporidií s houbami zprvu mluvily jen podobnosti neobvyklého průběhu meiotického cyklu mikrosporidií a některých hub, ale nyní je tato domněnka posílena fylogenezí bílkovin α a β tubulinu, jimiž se mikrosporidie řadí do fylogenetické radiace hub. Další podporou je, že ve fylogenetických rodokmenech se sekvence genů cpn70 větví, i když slabě, spolu se sekvencemi mitochondriálních genů hub rodu Boveria. Mikrosporidie tedy s velkou pravděpodobností nejsou potomci prastarých nebo primitivních eukaryontů, ve velké míře pozměněných adaptacemi k parazitizmu, ale naopak vzešly z organizmů značně pokročilých.
{kind=link}
Dvě základní charakteristiky archezoí, jejich primitivnost, místo na samém počátku vývoje a s tím spojená původní nepřítomnost mitochondrií, byly tedy zpochybněny. Ukazuje se, že i dva základní sloupy archezoí, metamonády a mikrosporidie, pokročily hodný kus cesty od uskutečnění mitochondriální symbiózy, ke které došlo daleko dříve než k jejich vzniku a patrně také dříve než k vzniku všech dnes známých eukaryont. Další existence zvláštní říše Archezoa je tedy neudržitelná.
Proč se mikrosporidie odvětvují na bázi eukaryont, jestliže vznikly z „terminální koruny“?
 Molekulární biologové na to mají různé odpovědi. Vysoce divergentní sekvence SSU rRNA se mohou umístit ve fylogenetických stromech na nesprávném místě, a v genech mikrosporidií je skutečně podíl substitucí výjimečně vysoký. Takové geny se někdy druží přednostně s jinými vysoce divergentními substitucemi, a to u fylogenetického stromu znamená velmi časné větvení. Dále, rodokmeny založené na podjednotkách ribozomální RNA jsou navzájem těžko slučitelné s těmi, které jsou založeny na genech kódujících bílkoviny, jako např. α a β tubulin. Řadu dalších léček uváději ve svých pracích A. Adoutte a H. Philippe. Podtrhují, že molekulární fylogeneze zpřesnila charakteristiku monofyletických skupin protistů, identifikovaných elektronovou mikroskopií, ale potýká se s podobnými potížemi jako elektronová mikroskopie, pokud chce přesně určit, v jakém pořadí skupiny vznikaly. To je způsobeno také tím, že soudobé formy patrně vznikly bouřlivým šířením (radiací) během relativně krátkého časového údobí.
Molekulární biologové na to mají různé odpovědi. Vysoce divergentní sekvence SSU rRNA se mohou umístit ve fylogenetických stromech na nesprávném místě, a v genech mikrosporidií je skutečně podíl substitucí výjimečně vysoký. Takové geny se někdy druží přednostně s jinými vysoce divergentními substitucemi, a to u fylogenetického stromu znamená velmi časné větvení. Dále, rodokmeny založené na podjednotkách ribozomální RNA jsou navzájem těžko slučitelné s těmi, které jsou založeny na genech kódujících bílkoviny, jako např. α a β tubulin. Řadu dalších léček uváději ve svých pracích A. Adoutte a H. Philippe. Podtrhují, že molekulární fylogeneze zpřesnila charakteristiku monofyletických skupin protistů, identifikovaných elektronovou mikroskopií, ale potýká se s podobnými potížemi jako elektronová mikroskopie, pokud chce přesně určit, v jakém pořadí skupiny vznikaly. To je způsobeno také tím, že soudobé formy patrně vznikly bouřlivým šířením (radiací) během relativně krátkého časového údobí.{kind=link}

 Základem této radiace byla snad událost mitochondriální symbiózy, ke které došlo patrně jen jedenkrát před miliardou let, když koncentrace kyslíku v atmosféře dosáhla zhruba dnešní hodnoty. Z tohoto předka pocházejí snad všichni existující protisti. S tím je spojena otázka vztahu mitochondrií a hydrogenozomů a příbuznosti protistů vybavených mitochondriemi s těmi, kteří je nemají, čili problém, který stál u zrodu archezoí. Novou hypotézou se na to r. 1998 pokusili odpovědět W. Martin a M. Müller. Podle obecně přijímané endosymbiotické teorie Lynn Margulisové dávná fagocytující archebakterie pohltila aerobní bakterii, patrně α-proteobakterii, ale nestrávila ji, podržela si ji jako symbionta. Ten dýchal, poskytoval ATP výměnou za metabolizovatelné substráty a úkryt, jeho geny byly z valné části přeneseny do genomu archebakterie, a tak se vyvinula mitochondrie. Martin a Müller naopak předpokládají, že autotrofní metanogenní archebakterie pohltila fermentující α-proteobakterii. Vyvinul se podobný vztah vzájemné závislosti, a zároveň přeměna hostitelské archebakterie z autotrofní na heterotrofní. Posléze se symbiont přeměnil v hydrogenozom, nebo, za podmínek globálního vzestupu hladiny atmosférického kyslíku a díky své dřívější metabolické rozrůzněnosti se vyvinul v mitochondrii, anebo se z buňky hostitele ztratil. Autoři nedávají tyto pochody do souvislostí se vznikem cytoskeletálních a endomembránových systémů, ale trvají na tom, že obě organely, mitochondrie i hydrogenozom, měly společného předka. Jako každá teorie bude i tato prověřována dalším výzkumem, ale zatím – podle názoru obou autorů – lépe vysvětluje energetický metabolizmus eukaryotických buněk bez mitochondrií.
Základem této radiace byla snad událost mitochondriální symbiózy, ke které došlo patrně jen jedenkrát před miliardou let, když koncentrace kyslíku v atmosféře dosáhla zhruba dnešní hodnoty. Z tohoto předka pocházejí snad všichni existující protisti. S tím je spojena otázka vztahu mitochondrií a hydrogenozomů a příbuznosti protistů vybavených mitochondriemi s těmi, kteří je nemají, čili problém, který stál u zrodu archezoí. Novou hypotézou se na to r. 1998 pokusili odpovědět W. Martin a M. Müller. Podle obecně přijímané endosymbiotické teorie Lynn Margulisové dávná fagocytující archebakterie pohltila aerobní bakterii, patrně α-proteobakterii, ale nestrávila ji, podržela si ji jako symbionta. Ten dýchal, poskytoval ATP výměnou za metabolizovatelné substráty a úkryt, jeho geny byly z valné části přeneseny do genomu archebakterie, a tak se vyvinula mitochondrie. Martin a Müller naopak předpokládají, že autotrofní metanogenní archebakterie pohltila fermentující α-proteobakterii. Vyvinul se podobný vztah vzájemné závislosti, a zároveň přeměna hostitelské archebakterie z autotrofní na heterotrofní. Posléze se symbiont přeměnil v hydrogenozom, nebo, za podmínek globálního vzestupu hladiny atmosférického kyslíku a díky své dřívější metabolické rozrůzněnosti se vyvinul v mitochondrii, anebo se z buňky hostitele ztratil. Autoři nedávají tyto pochody do souvislostí se vznikem cytoskeletálních a endomembránových systémů, ale trvají na tom, že obě organely, mitochondrie i hydrogenozom, měly společného předka. Jako každá teorie bude i tato prověřována dalším výzkumem, ale zatím – podle názoru obou autorů – lépe vysvětluje energetický metabolizmus eukaryotických buněk bez mitochondrií.
Literatura
P. J. Keeling: A kingdom´s progress: Archezoa and the origin of eukaryotes, BioEssays 20, 87–95, 1998W. Martin a M. Müller: The hydrogen hypothesis for the first eukaryote, Nature 392, 37–41, 1998
H. Philippe a A. Adoutte (1996): How far can we trust the molecular phylogeny of protists? Ver. Deutsch. Zool. Ges. 89, 49–62, 1996/2
Ke stažení
 Článek ve formátu PDF [947,53 kB]
Článek ve formátu PDF [947,53 kB]
O autorovi
Jiří Lom
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
