









Životně důležité tance
Existují biologické zákonitosti, o kterých se ve slušné společnosti nepochybuje. Patří k nim i tvrzení, že buňky vznikají výhradně dělením existujících buněk. Protože se buňky dovedou dělit už od pradávna, lze očekávat, že sama procedura dělení je záležitost evolučně stará a příslušně konzervativní. Na první pohled se zdá oprávněnou domněnka, že – právě tak jako genetický kód – i buněčné dělení je jenom jedno.
 Při dělení buněk se zejména pečlivě dělí jejich genetický materiál, aby obě vzniklé dceřiné buňky získaly stejnou sadu instrukcí, kódovaných v molekulách DNA.
Při dělení buněk se zejména pečlivě dělí jejich genetický materiál, aby obě vzniklé dceřiné buňky získaly stejnou sadu instrukcí, kódovaných v molekulách DNA.  Tyto instrukce (genom) určují strukturu a tvar buněk, jejich biochemické vybavení, popřípadě jejich místo a osud v mnohobuněčném organizmu. Každá buňka našeho těla potřebuje k uložení příslušných informací asi tak metr molekul DNA, které se musí v buňce pečlivě poskládat, aby se tam vešly, a před každým dělením zkopírovat.
Tyto instrukce (genom) určují strukturu a tvar buněk, jejich biochemické vybavení, popřípadě jejich místo a osud v mnohobuněčném organizmu. Každá buňka našeho těla potřebuje k uložení příslušných informací asi tak metr molekul DNA, které se musí v buňce pečlivě poskládat, aby se tam vešly, a před každým dělením zkopírovat.  To samo o sobě předpokládá dokonalost prostorového uspořádání, která je na samé hranici pochopitelnosti. A což teprve neomylně přesné rozdělování kopií genomu mezi dceřiné buňky při dělení, zvlášť když si uvědomíme, že naše metrová molekula je rozdělena na 46 různých úseků – chromozomů ! A jsme na tom ještě dobře ve srovnání s kaprem, který má 104 chromozomů, kdežto naopak komárovi jich stačí šest.
To samo o sobě předpokládá dokonalost prostorového uspořádání, která je na samé hranici pochopitelnosti. A což teprve neomylně přesné rozdělování kopií genomu mezi dceřiné buňky při dělení, zvlášť když si uvědomíme, že naše metrová molekula je rozdělena na 46 různých úseků – chromozomů ! A jsme na tom ještě dobře ve srovnání s kaprem, který má 104 chromozomů, kdežto naopak komárovi jich stačí šest.
Důmyslný tanec chromozomů
Proč se chromozomům říká chromozomy čili „barevná tělíska“? Nejspíš proto, že v době, kdy byly poprvé pozorovány, nikdo nevěděl, k čemu jsou dobré; do objevu genetické funkce DNA zbývalo ještě třičtvrtě století. Jedinou jejich nepochybnou vlastností bylo, že se při určitých histologických reakcích barvily. Spojení mezi chromozomy a dědičností vytušil koncem minulého století A. Weismann, ten, který stříhal ocásky myším, aby ukázal, že bezocasost není dědičná (nezaměňovat s jistým sovětským badatelem z poloviny našeho století, který prý dělal totéž, aby ukázal, že bezocasost dědičná je). Napadlo ho totiž, že rozdělování „zárodečné plazmy“ (dnešním slovníkem: nositele dědičné informace) by mohlo být účelem podivného chování chromozomů, které očividně v dělících se buňkách vytvářely složité „taneční figury“. Jediným zjevným cílem chromozomálního tance bylo „rozpočítání“ chromozomů na dvě skupiny, jež se pak rozcházely do vznikajících dceřiných jader. Pro celý proces se vžilo označení mitóza, ve starší české literatuře se setkáváme s názvem „nepřímé dělení jaderné“ (viz obrázek a obrázek a snímky J. Pláška – obrázek).Genom se dá dělit všelijak
Staromódní české označení mitózy naznačuje, že komplikovaný chromozomální tanec nemusí být obecnou součástí života všech eukaryontních buněk. Přinejmenším by mohlo existovat jakési „přímé“ dělení, které by dosahovalo téhož cíle – rozdělení jádra na stejné části – jednodušším způsobem. Pozorování některých buněk ve světelném mikroskopu tuto možnost zdánlivě potvrzuje. U kvasinek se jádro, bez ohledu na postup buněčného dělení, jeví jako jednolitá hmota, která se v příslušném okamžiku prostě přeškrtí na dvě poloviny. Někteří cytologové minulosti se odvažovali hovořit o tom, že kvasinky provozují amitózu čili přímé dělení jaderné; jiní usilovně zdokonalovali mikroskopické techniky, aby dokázali, že kvasinka provádí docela normální mitózu s docela normálními chromozomy. Pozdější velký klasik kvasinkové genetiky, Carl C. Lindegren, dokonce ve 40. a 50. letech publikoval údajné pozorování kvasinkových chromozomů a mitotických figur. Jak vidět, i klasikové se mohou mýlit – struktury, které Lindegren pokládal za chromozomy, byly zřejmě arteficiální sraženiny ve vakuole. Věrohodný popis jaderného dělení u kvasinek umožnila až kombinace technik elektronové a světelné mikroskopie a genetických pozorování; i ukázalo se, že – jako obvykle – v obou protichůdných názorech lze nalézt kousek pravdy. Kvasinky totiž provozují zvláštní variantu nepřímého dělení jaderného, označovanou jako endomitóza; je to verze klasické mitózy, která se odehrává uvnitř neporušené blány jaderné. Navíc kvasinkové chromozomy zůstávají rozvolněné i v průběhu dělení jádra, takže je nelze přímo pozorovat. Chybějí zde tedy dva základní rysy „klasické“ mitózy – a my jsme o zkušenost bohatší. Víme totiž, že ani rozpad jaderné blány, ani kondenzace chromozomů nejsou pro pravidelný průběh jaderného dělení nezbytné.Dalším bohatým zdrojem zvláštních případů je zřejmě jaderné dělení dalších jednobuněčných organizmů, zejména prvoků. Prvoci označovaní jako Dinokaryota („podivnojaderní“) mají chromozomy kondenzované i v interfázi; struktura jádra jiných skupin (či spíše jejich „dvojjádří“ složené z mikronukleu a makronukleu) naznačuje, že ani tady s obyčejnou mitózou nevystačíme. Dokonce i u mnohobuněčných živočichů se můžeme setkat s neobvyklým uspořádáním chromozomů – někteří členovci mají tzv. holocentrické či holokinetické chromozomy – tedy chromozomy bez centromery či spíše chromozomy, které jsou celé samy sobě centromerou.
Nezdá se však, že by nějaký organizmus skutečně provozoval amitózu. Ta byla s postupem doby odsunuta téměř do zapomenutí, i když se o ní dodnes čas od času diskutuje (viz text v rámečku).
Meióza aneb pánové, zadejte si
Ani „velcí eukaryonti“, jako jsme my, nevystačí po celý život s jediným způsobem jaderného dělení dle učebnicového schématu mitózy. Pohlavně se rozmnožující organizmy mohou dosáhnout shody v obecném uspořádání genomu mezi po sobě následujícími generacemi jen tehdy, když jednou za životní cyklus – v průběhu tvorby pohlavních buněk (gamet) – sníží počet chromozomů na polovinu. Pomineme-li pro jednoduchost rozdíly v počtu a struktuře chromozomů jednotlivých pohlaví, je genotyp jedince zdvojený (diploidní), zatímco genotyp jeho gamet je jednoduchý (haploidní): buňky tkání našeho těla mají 46 chromozomů, vajíčka a spermie 23. Haploidní buňky vznikají z diploidních redukčním dělením čili meiózou (obrázek).Proces meiózy se v mnohém podobá mitóze; meiotické dělení však před buňku staví ještě jeden úkol, se kterým se mitotická buňka nesetká. Podmínkou úspěšného redukčního dělení je, že se sobě odpovídající (strukturně shodné) chromozomy navzájem najdou. Není to úkol jednoduchý – ani příroda se nedokáže vždycky vyvarovat omylů. Mnohé genetické vady (z těch lidských je asi nejznámější Downův syndrom čili trisomie – ztrojení – chromozomu 21) jsou právě důsledkem chyb při meióze.
Opravdové nesnáze nastávají u organizmů, jejichž genom se meiózou dělí na dvě nestejně velké skupiny chromozomů. Není jich mnoho; patří k nim například některé skupiny hmyzu, u nichž je pohlaví určeno lichým počtem chromozomů. „Spořádaní“ živočichové, jako jsme my savci, vystačí s jedním párem pohlavních chromozomů. Jedno pohlaví má oba pohlavní chromozomy stejné, produkuje jediný typ gamet a nechává určení pohlaví potomstva na partnerovi; partner má pohlavní chromozomy různé a produkuje dvojí gamety, z jedněch jsou pak po oplození dcery, z druhých synové. 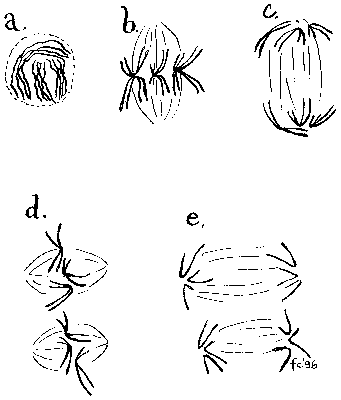 U některých členovců je však určení pohlaví podstatně složitější: ještě poměrně jednoduchá je situace u kudlanky nábožné, jejíž sameček se při meióze musí postarat o tři pohlavní chromozomy (obrázek). Uspořádat a rozdělit trojici pohlavních chromozomů dá více práce než rozdělit pár; zhruba v každé desáté buňce se připojení třetího pohlavního chromozomu k mikrotubulům vřeténka nezdaří. Kudlanka se vyhýbá z toho pramenícím potížím tím, že postiženým buňkám nedovolí dokončit mitózu a dozrát ve spermie, pokud se „ztracený“ chromozom nepřichytí k vřeténku.
U některých členovců je však určení pohlaví podstatně složitější: ještě poměrně jednoduchá je situace u kudlanky nábožné, jejíž sameček se při meióze musí postarat o tři pohlavní chromozomy (obrázek). Uspořádat a rozdělit trojici pohlavních chromozomů dá více práce než rozdělit pár; zhruba v každé desáté buňce se připojení třetího pohlavního chromozomu k mikrotubulům vřeténka nezdaří. Kudlanka se vyhýbá z toho pramenícím potížím tím, že postiženým buňkám nedovolí dokončit mitózu a dozrát ve spermie, pokud se „ztracený“ chromozom nepřichytí k vřeténku.  Organizmus tedy musí udržovat přehled o osudu jednotlivých chromozomů; to jsme ostatně mohli tušit už podle obdivuhodné spolehlivosti meiotického (i mitotického) procesu. Zvláštní uspořádání chromozomů a neobvykle velké buňky kudlanky nábožné pak umožnily přejít od tušení k exaktnímu studiu mechanizmů, které „hlídají“ přichycení chromozomů k vřeténku. Mikrochirurgické pokusy ukázaly, že buňky vnímají mechanické napětí chromozomů – zataháme-li za nepřichycený chromozom mikrojehlou, meióza pokračuje, jako by vše bylo v pořádku! Toto pozorování je možná prvním podstatným krokem, který otvírá cestu k hledání možných mechanoreceptorů na chromozomech. Dobrými kandidáty by mohly být třeba nedávno popsané chromozomální proteiny, jejichž barvitelnost pomocí protilátek závisí na tom, zda je chromozom připojen k vřeténku.
Organizmus tedy musí udržovat přehled o osudu jednotlivých chromozomů; to jsme ostatně mohli tušit už podle obdivuhodné spolehlivosti meiotického (i mitotického) procesu. Zvláštní uspořádání chromozomů a neobvykle velké buňky kudlanky nábožné pak umožnily přejít od tušení k exaktnímu studiu mechanizmů, které „hlídají“ přichycení chromozomů k vřeténku. Mikrochirurgické pokusy ukázaly, že buňky vnímají mechanické napětí chromozomů – zataháme-li za nepřichycený chromozom mikrojehlou, meióza pokračuje, jako by vše bylo v pořádku! Toto pozorování je možná prvním podstatným krokem, který otvírá cestu k hledání možných mechanoreceptorů na chromozomech. Dobrými kandidáty by mohly být třeba nedávno popsané chromozomální proteiny, jejichž barvitelnost pomocí protilátek závisí na tom, zda je chromozom připojen k vřeténku.
A ponaučení?
Někdy jsme v pokušení zanedbávat netypické situace, zvláštní případy a výjimky z pravidel, a namlouváme si, že organizmy se ve všech základních buněčných funkcích chovají v podstatě stejně. Není tomu tak – dokonce ani v případě funkce tak podstatné, jako je zajištění rovnoměrného rozdělování genetické informace mezi dceřiné buňky. A co víc – kdyby tomu tak bylo, přišli bychom o příležitost pochopit mnohé o skutečně obecných mechanizmech jaderného dělení, které můžeme studovat právě díky netypickým situacím, zvláštním případům a výjimkám z pravidel!Literatura
K. Hrubý: Genetika. SPN, Praha 1961R. K. Mortimer: Carl C. Lindegren: the iconoclastic father of Neurospora and yeast genetics. The Early Days of Yeast Genetics, Cold Spring Harbor Laboratory Press, Cold Spring Harbor 1993, pp. 17–38
J. Král: Holokinetické chromozómy. Biol. Listy 59,191–217, 1994
X. Li a R. B. Nicklas: Mitotic forces control a cell-cycle checkpoint. Nature 373, 630–632, 1995
Obrázky
Poznámky
Existuje amitóza, či ne?
Dělení buněčného jádra mitózou představuje mechanizmus, kterým dvě dceřiné buňky dostávají stejný podíl genetické informace. Při mitóze dochází k nápadným morfologickým změnám, jež jsou dobře známé: patrně všichni biologové, nejenom cytologové, někdy v mikroskopu viděli mitotické figury s chromozomy (viz obrázek). Starší učebnice biologie však obvykle obsahují zmínku, že vedle mitózy (nepřímého dělení) se jádra eukaryotických buněk mohou dělit amitózou (přímým dělením). Při amitóze chromozomy nevznikají, jaderná hmota se dělí vchlípením jaderné membrány a zaškrcením jádra. Posledních asi dvacet let se spíše soudí, že amitóza u buněk mnohobuněčných organizmů se za normálních životních podmínek nevyskytuje. Jen málokterý biolog totiž tvrdí, že amitózu pozoroval. Ale ojediněle se objevují publikace, jejichž autoři takové tvrzení uvádějí.
Patrně poslední takovou publikací je práce S. Barniho a spol. z Padovské univerzity (Comp. Haematol. Int. 5, 183 188, 1995), v níž jsou otištěny výsledky pozorování preparátů krvetvorné tkáně jater mloků v optickém i elektronovém mikroskopu. Autoři uvádějí, že v této tkáni se krvetvorné buňky dělí pouze amitoticky. Svá pozorování doporovázejí velmi kvalitními mikrofotografiemi, na kterých je naprosto zřetelné zaškrcení jádra nebo výskyt dvou jader v jedné buňce.
Podotkněme, že ale v bílých krvinkách savců (s nimiž Barniho skupina své nálezy neporovnává) se ojediněle vyskytují vedle jádra rovněž úlomky jaderné hmoty. Takto se v savčích bílých krvinkách vyskytují dva útvary s jadernou hmotou, přičemž vznikají za patologických podmínek. Uvedený úkaz lze u savců experimentálně navodit podáním některých hemotoxických xenobiotik. Ze stejného důvodu se vyskytují podobné úlomky jaderné hmoty i v erytroblastech a zůstávají přítomny i v bezjaderných červených krvinkách, které z erytroblastů vznikají. Je na tom založen jeden ze starších testů na sledování mutagenity látek (tzv. mikronukleus test).
Takové nálezy v savčích krvinkách samozřejmě nejsou důkazem neexistence amitózy. Nicméně ukazují, že se jádro může rozdělit na dvě a více částí, ty mohou dále existovat, aniž to má přirozený funkční význam, který bychom od amitózy očekávali (tak jako amputaci končetiny při úrazu nepovažujeme za rozmnožování). Dokonce i zaškrcování buněčného jádra se běžně u části zralých savčích krvinek přirozeně vyskytuje (tzv. segmentované jádro granulocytů), aniž by následovalo jaderné dělení. Takže ani zaškrcení jádra nemůže být důkazem následné amitózy.
Zdá se, že existence amitózy u mnohobuněčných organizmů žijících v obvyklých životních podmínkách stále hodnověrně doložena není. Kdyby někdo popsal základní mechanizmus amitózy (jako je tomu dlouhá léty u mitózy), nebylo by pochyb. Ale samotné občasné fotografie zaškrceného jádra neumožňují, aby z nich vyplývající úvahy o možné existenci amitózy u mnohobuněčných organizmů opustily oblast nedostatečně doložených domněnek.
Kinetochory, mitotické vřeténko a pohyb chromozomů
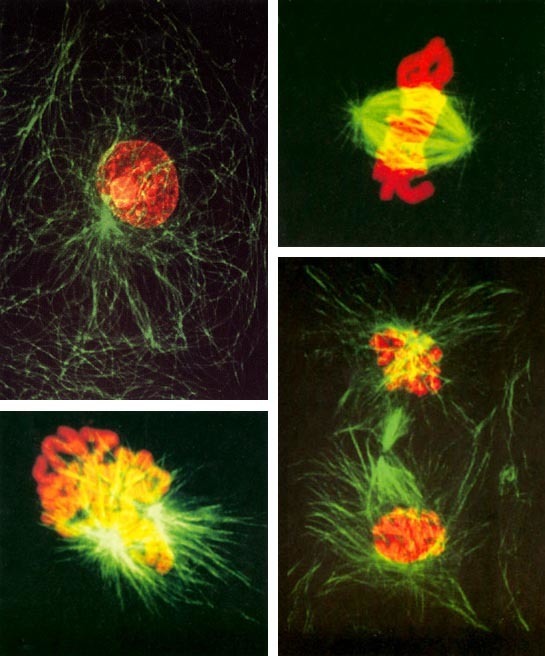
Poznámka k barevným obrázkům
Při nepřímém dělení buňky, mitóze, se v prometafázi (počátku druhé fáze mitózy) vytváří mitotické vřeténko, jeho vlákna se skládají z mikrotubulů, jejich stavebním materiálem jsou polymerizované cytoskeletové proteiny, tubulin α a tubulin β. Mikrotubuly jsou jedním svým koncem vázány na centriol, druhým na tzv. centromer (neboli kinetochor) některého z čerstvě rozdělených chromozomů. Připojením mikrotubulů ke kinetochorům je odstartována třetí fáze mitózy, anafáze, během ní se od sebe vzdalují sesterské chromozomy. Silou vyvíjenou zkracujícími se mikrotubuly jsou tyto chromozomy taženy k centriolům, nacházejícím se v protilehlých pólech mitotického vřeténka.
Mechanizmus mitózy dosud zůstává jedním ze základních a ne zcela dořešených problémů biologie. Je zřejmé, že jedním z nevyhnutelných kroků na cestě k objasnění tohoto jevu je podrobné studium chování chromozomů v různých fázích mitózy. V poslední době začala být k tomuto účelu využívána též konfokální fluorescenční mikroskopie (Vesmír 74, 508, 1995/9). Pomocí vhodných fluorescenčních sond a značek lze selektivně zviditelňovat chromozomy i vlákna mitotického vřeténka. V kombinaci s možností zobrazovat optické řezy buňkou, které lze postupně snímat z různé hloubky, umožňuje pak konfokální fluorescenční mikroskopie získat představu o vzájemném prostorovém uspořádání chromozomů a vláken mitotického vřeténka. Na rozdíl od elektronové mikroskopie však takovou informaci získáme, aniž bychom buňku museli fyzicky rozřezat na tenké plátky, což znamená, že na jedné vybrané buňce můžeme studovat i kinetiku chromozomálních pohybů.
O autorovi
Fatima Cvrčková

Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
