









Blížíme se k poznání podstaty biologických hodin?
| 5. 8. 1994Všechny živé organizmy, od nejjednodušších po nejsložitější, vykazují denní rytmy. Například u jednobuněčné mořské řasy Gonyaulax polyedra lze vysledovat, mimo jiné, denní rytmus v záření: v určitou noční dobu se oceán u kalifornského pobřeží, kde řasa žije, rozzáří světélkováním tisíců řas. Nejsložitější živý organizmus, člověk, dodržuje pravidelný rytmus spánku a bdění, kolísání tělesné teploty a četné jiné. Co mají všechny tyto denní rytmy v živých organizmech společného?
Předně jsou to rytmy vnitřní, endogenní. Přetrvávají i ve zcela neperiodických prostředích, např. ve stálé tmě bez jakéhokoliv signálu o vnějším času. Protože jsou endogenní, musí být poháněny vnitřními hodinami, pacemakerem. Tento pacemaker v neperiodickém prostředí neběží s periodou přesně 24hodinovou, ale udržuje ji jen zhruba (cirka); proto jsou hodiny i jimi řízené rytmy nazývány cirkadiánní. S 24hodinovým dnem je cirkadiánní pacemaker synchronizován převážně pravidelným střídáním světla a tmy. Je to světlá část dne, která hodiny synchronizuje. Je-li vnitřní perioda hodin kratší než 24 h, je pro synchronizaci hodin s 24hodinovým dnem důležitý konec světlé části dne při soumraku, který hodiny zpožďuje. Je-li naopak perioda hodin delší než 24 h, bude pro jejich synchronizaci s vnějším dnem podstatné první světlo při ranním úsvitu, které indukuje jejich předbíhání.
Cirkadiánní vnitřní hodiny
Aby cirkadiánní pacemaker mohl plnit funkci vnitřních biologických hodin, musí splňovat určité podmínky. Jsou-li orgán či buňka, o nichž předpokládáme, že obsahují pacemaker, chovány v živném roztoku vně organizmu, musí vykazovat sebeudržující, zhruba 24hodinové oscilace, např. v elektrické aktivitě, metabolizmu, tvorbě různých látek apod. Pacemaker musí být teplotně kompenzován, neboli rychlost chodu hodin nesmí záležet na malých teplotních výkyvech. Je-li pacemaker v organizmu zničen, organizmus se stane arytmickým, neprojevují se u něj denní rytmy. Je-li takto arytmickému organizmu zpětně implantován pacemaker z cizího dárce, může být denní rytmicita organizmu obnovena. Tomuto popisu vlastností pacemakeru vyhovují cirkadiánní hodiny uložené v očích mořského měkkýše Aplysia, či pacemaker uložený v epifýze vrabce domácího, nebo nakonec i hodiny v části mozku savců, a tedy i člověka, zvané hypotalamus; nositeli oscilací v hypotalamu jsou dvě suprachiazmatická jádra, což jsou shluky neuronů uložené poblíž optického chiazmatu (křížení) po obou stranách třetí komory mozkové.Přesto, že byly částečně popsány formální vlastnosti biologických hodin, jejich synchronizace a umístění v organizmech, dosud málo toho bylo řečeno o pravé podstatě denních oscilací v hodinách. Bylo zřejmé, že podkladem cyklických změn by mohl být zpětný, seberegulující, zhruba 24hodinový cyklicky se opakující proces, ale nedařilo se tento problém rozřešit. Teprve užití metod molekulární biologie umožnilo v poslední době přiblížit se k podstatě oscilací. Otázka se řešila převážně na dvou jednodušších modelech: ovocnou muškou druhu Drosofila melanogaster se zabývali vědci shromáždění kolem J. C. Halla a M. Rosbashe na Brandeisově univerzitě a kolem M. W. Younga na Rockefellerově univerzitě, a chlebovou plísní druhu Neurospora crassa vědci, které kolem sebe shromáždili J. C. Dunlap a J. J. Loros na Dartmouthské lékařské škole. Výsledky všech škol jsou si vzácně blízké a byla jim v posledních letech, zvláště pak letos, věnována velká pozornost na stránkách významných časopisů, např. Science.
Cirkadiánní rytmy ovocné mušky
U jednotlivých mušek lze pozorovat denní rytmy v pohybové aktivitě: jsou aktivní v době svého subjektivního dne, a to i v neperiodickém prostředí. U populace mušek byly zaznamenány cirkadiánní rytmy v líhnutí z kukel: líhnou se vždy v časných ranních hodinách. Za oba tyto rytmy je zodpovědný jeden gen, tj. část deoxyribonukleové kyseliny (DNA) v jádru buňky. Tento gen se nazývá „per“, neboť kóduje tvorbu produktu podmiňujícího opakující se děje o periodě přibližně 24 h. Chemickou cestou či ozařováním byly připraveny cirkadiánní mutanty mušky, tj. mušky se změněnou periodicitou: perO, perS a perl. Muška s genem perO je arytmická: nebyly u ní pozorovány denní rytmy ani v pohybové aktivitě, ani v líhnutí. Je-li však do zárodečné buňky této mušky vpraven gen per z divokého, rytmického kmene, bude se muška s původním genem perO chovat též rytmicky. Mušky s genem perS mají periodu endogenních hodin pouze 19hodinovou (s je z anglického short, tj. krátký), mušky s genem perl mají periodu 29hodinovou (l je z anglického long, tj. dlouhý). Gen per byl již izolován a pořadí bází v DNA, čili sled „kódujících písmen“, bylo stanoveno. Gen kóduje per-protein, který obsahuje zhruba 1 200 aminokyselin. Bodová mutace jedné jediné báze a následná změna jedné z aminokyselin v per-proteinu vede k výraznému zkrácení (perS), nebo naopak k prodloužení (perl) vnitřní periody.Zpětnovazebná smyčka tvorby per-proteinu
Per-protein se tvoří rytmicky v hlavě ovocné mušky. Maximálních koncentrací dosahuje v pozdní subjektivní noci, tj. k ránu. U arytmické mušky s mutací perO se hladina proteinu cyklicky nemění. U mušky s genem perS se hladina proteinu mění s periodou 19 h, u mušky s perl s periodou 29 h; perioda cyklické tvorby per-proteinu tedy odpovídá periodě rytmu v pohybové aktivitě či líhnutí. Cyklickým změnám v hladině per-proteinu předcházejí cyklické změny v množství odpovídající informační ribonukleové kyseliny (per-mRNA; m z angl. messenger – posel). Tato mRNA se přepisuje v jádře buňky z DNA per-genu. Na ní je pak jako na matrici tvořen per-protein. Per-mRNA dosahuje maxima v časných nočních hodinách, o 6 – 8 hodin dříve než vlastní per-protein. Vzhledem k tomu, že per-protein je v hlavě mušky umístěn v jádrech nervových buněk a že jde tedy o jaderný protein, nabízela se představa, že by mohl v jádře buňky ovlivňovat přepis své vlastní mRNA z per-genu.Smyčka by potom vypadala takto: na per-genu (DNA) se začíná přepisovat mRNA pro tvorbu per-proteinu, vzniká a hromadí se per-mRNA, na ní se překládá per-protein, hromadí se a při větším množství začíná blokovat (inhibovat) přepis per-mRNA z per-genu – množství per-mRNA klesá a v důsledku toho klesá i množství per-proteinu atd. Tato jednoduchá představa zpětné smyčky byla svůdná, jejímu přijetí však bránila skutečnost, že per-protein se na DNA neváže a nemůže tudíž sloužit jako přímý regulační faktor.
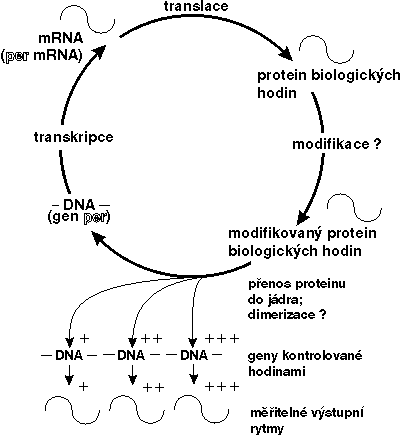
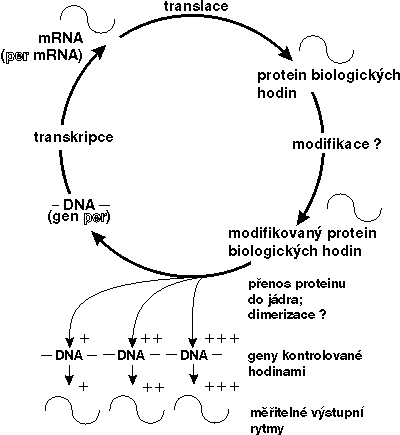 Výzkumy v posledních dvou letech však ukázaly, že per-protein by přesto regulačním faktorem být mohl a mohl by i ovlivňovat přepis vlastní mRNA. Per-protein totiž obsahuje ve své molekule oblast zvanou PAS, kterou se může vázat s jinými proteiny obsahujícími obdobnou oblast a tvořit s nimi dimery (tj. sloučeniny vzniklé spojením dvou molekul monomeru). Tyto jiné proteiny pak mohou mít schopnost vázat se na DNA v jádře a ovlivňovat transkripci. Jako dimer může být zřejmě per-protein navázán na per-gen. A zřejmě nejen na něj. Jako transkripční faktor by mohl ovlivňovat přepis více genů řízených cirkadiánními hodinami „dolů po proudu“ od hodin centrálních (viz obrázek).
Výzkumy v posledních dvou letech však ukázaly, že per-protein by přesto regulačním faktorem být mohl a mohl by i ovlivňovat přepis vlastní mRNA. Per-protein totiž obsahuje ve své molekule oblast zvanou PAS, kterou se může vázat s jinými proteiny obsahujícími obdobnou oblast a tvořit s nimi dimery (tj. sloučeniny vzniklé spojením dvou molekul monomeru). Tyto jiné proteiny pak mohou mít schopnost vázat se na DNA v jádře a ovlivňovat transkripci. Jako dimer může být zřejmě per-protein navázán na per-gen. A zřejmě nejen na něj. Jako transkripční faktor by mohl ovlivňovat přepis více genů řízených cirkadiánními hodinami „dolů po proudu“ od hodin centrálních (viz obrázek).
{kind=link}
Smyčka tedy byla navržena. Je vysvětlení tak lehké? Ve hře je ještě mnoho neznámých. Proč je tak dlouhá doba mezi nahromaděním transkriptu per-mRNA a maximální hladinou per-proteinu? Je per-protein v dimerizované podobě již vlastním transkripčním faktorem, či musí ještě projít posttranslačními obměnami, např. fosforylací? Jak rychle se per-protein či jeho meziprodukt rozkládá? Otázek je zatím více než zjištěných skutečností, ale základ naší představy sebeudržujících oscilací byl již položen.
V poslední době byly popsány další cirkadiánní mutanty u mušky druhu Drosophila melanogaster, tzv. tim (z anglického timeless – bez času). Tyto mutanty jsou skutečně bez časového řádu, arytmické. Zdá se, že jejich per-proteiny nejsou schopny se vázat v jádře nervových buněk. Nemohou tak plnit úlohu transkripčního faktoru, který inhibuje přepis své vlastní per-mRNA.
Nastavování biologických hodin per-proteinem
Je-li per-protein a jeho tvorba součástí cirkadiánního pacemakeru, a tedy jeho zpětné smyčky, mělo by umělé zvýšení tvorby per-proteinu v neodpovídající době vést k fázovému posunu pacemakeru a k jeho přesunu do jiného vnitřního času. K tomu také skutečně dochází. Vpraví-li se do zárodku ovocné mušky takový per-gen, který umožní zahájit tvorbu per-proteinu pomocí teplotního podnětu, pak v závislosti na době působení tohoto podnětu může dojít nejen k prudkému vzrůstu tvorby per-proteinu, ale současně i k fázovému posunu, jmenovitě k předběhnutí rytmu v pohybové aktivitě. Změna koncentrace per-proteinu tedy může vyvolat i změnu fáze hodin.Rytmický přepis mRNA, analogické per-mRNA ovocné mušky, byl nalezen i v biologických hodinách jiných organizmů, např. v suprachiazmatických jádrech potkana. Tato mRNA se ve větším množství přepisuje z DNA v době subjektivního dne než v době subjektivní noci. Je-li však potkan v noci osvícen, světelný podnět okamžitě fázově posune cirkadiánní pacemaker do jiného času a dojde k prudkému vzestupu transkripce. I v cirkadiánním pacemakeru savců dochází tedy v okamžiku fázového posunu ke změnám v této mRNA, která může být tudíž i součástí hodin savců.
Biologické hodiny v chlebové plísni
Chlebová plíseň druhu Neurospora crassa je vláknitá houba, v níž biologické hodiny řídí rytmus cyklické produkce asexuálních spor, tzv. konidií. Součástí hodin je gen „frq“ (z angl. frequency – frekvence cyklu = 1/perioda cyklu). Obdobně jako u drosofily, byly i u neurospory izolovány cirkadiánní mutanty s periodou výrazně kratší nebo výrazně delší než 24 hodin a mutanty zcela arytmické. Gen frq kóduje tvorbu proteinu FRQ, obdobně jako per-gen u mušky kóduje tvorbu per-proteinu. Zdá se, že zpětnovazebná smyčka u neurospory je velmi podobná smyčce u drosofily. Na genu frq vzniká frq-mRNA, jejíž rytmický přepis byl již prokázán, a frq-mRNA je překládaná do FRQ-proteinu. Tento protein může tlumit další tvorbu frq-mRNA. Tlumivý vliv FRQ-proteinu na transkripci vlasní mRNA byl již také prokázán. Sled kódujících bází je však odlišný u per a frq genu. Zpětnovazebná smyčka tvorby proteinu jako součásti biologických hodin se nicméně zdá být obdobná. FRQ-protein, obdobně jako per-protein, by mohl ovlivňovat nejen transkripci vlastního genu, ale též transkripci dalších genů „dolů po proudu“, tzn. genů kontrolovaných hodinami, jejichž rytmický přepis indukuje další cyklické změny.Cyklická tvorba proteinu: podstata biologických hodin?
Z popisu oscilací tvorby centrální bílkoviny u druhů Drosofila melanogaster a Neurospora crassa vyplývá, že zpětnovazebné ovlivnění této tvorby samotným proteinem by mohlo být podstatou biologických hodin řídících cirkadiánní rytmy v organizmech. Centrální protein by sloužil jako transkripční faktor řídící nejen přepis svého vlastního genu, ale i přepis dalších genů kontrolovaných hodinami. Model však zřejmě nebude tak jednoduchý. Je nutné uvažovat více neznámých: vazbu transkripčního faktoru na DNA; interval mezi přepisem do mRNA a tvorbou bílkoviny; posttranslační úpravy proteinu; účast dalších proteinů na transkripci; společnou sehranost těchto složek; účast dalších složek na zpětnovazebné smyčce; poločas rozpadu centrálního proteinu či jiných proteinů; poločas rozpadu mRNA a četné jiné.Dosud byly také popsány molekulární pochody v biologických hodinách pouze u dvou organizmů. Je tento model obecný? Jsou centrální proteiny analogické? Byl mechanizmus či gen biologických hodin konzervován během fylogeneze, tj. vývoje druhů? Tyto otázky mohou být opět řešeny metodami molekulární biologie. J. Takahashi z Centra pro biologické časování, které vytvořila Národní nadace pro vědu ve Spojených státech, již se svými kolegy přistoupil k analýze cirkadiánních hodin savců. Vyšlechtil cirkadiánní mutanty myši domácí s periodou rytmů pouhých 20 hodin. Vzhledem k tomu, že složení genomu myši je již známé, je možné hledat, který gen či která část DNA je změněna a odpovídá za drastické zkrácení periody.
Je možné však zvažovat i otázku, kdy a jak se v organizmech vytvořila a konzervovala zhruba 24hodinová zpětná smyčka? Smyčka o této délce je obdivuhodný přírodní výtvor. Je-li však čas měřen cyklicky touto smyčkou, může se jeho stále se opakující průběh jevit nevzrušivý: nárůst proteinu, pokles proteinu, nárůst proteinu atd.
Cyklické měření času tedy začíná být postupně odhalováno. Jen pomalu počíná být též odkrýváno tajemství dlouhodobého programu vývoje od zárodku ke smrti, a to opět převážně u ovocné mušky.
Tento neopakovatelný tok času popisuje Rainer Maria Rilke ve své básni
Konec podzimu:
Teď mi čas dokázal,
kterak se všechno mění.
Cos vyvstává a plení
a dělá zlo a žal.
Jak dny jdou dál,
žádný sad stejný není;
od žloutnoucích k těm žlutým,
pozvolna uchřadnutým
předlouho jsem se bral.
Až se lidé více přiblíží i k řešení tohoto dlouhodobého programu, budou o maličko blíže porozumění neopakovatelnému biologickému toku času. Nyní získávané poznatky o molekulární podstatě biologických hodin nám umožňují přiblížit se pochopení cyklického biologického toku času.
O autorovi
Helena Illnerová
Doporučujeme

Když bahno teče jako ledovec

Ideologie v mapách, mapy v rukách ideologů 
