










Vliv taxonomické metody na permské vymírání
Titulek článku zní tak trochu jako nesmysl, vždyť jaký by naše věda mohla mít vliv na katastrofu, která se udála hluboko v minulosti. Jenomže existují dvě permské události – ta skutečná, ke které došlo před miliony let a na kterou už opravdu nemůžeme mít vliv, a pak permská událost jako přírodovědecký femoném, jako soubor hypotéz, které nyní tvoříme svým vědeckým myšlenkovým aparátem a zakládáme na nepatrných štřípcích sporných faktů pocházejících ze skutečné minulosti. Ta druhá permská událost si ve vědecké komunitě žije svým vlastním životem a naše poznávací metody na ni určitě vliv mají.
Někdy se zdá, že celé bádání o minulosti života je jen chabě podloženou hrou s čísly. Tak je tomu i v knize známého evolučního biologa Davida M. Raupa „O zániku druhů“, která vyšla i u nás (viz recenzi ve Vesmíru 73, 456, 1994/8), a vlastně i v následujícím článku.
Na straně 65 a předešlých stranách se Raup zabývá hromadným vymíráním na hranici permu a triasu asi před 245 miliony let, největší známou katastrofou, která postihla pozemský život (blíže k tématu velkých vymírání viz např. Vesmír 73, 435, 1994/8). Podle Raupova odhadu při ní vyhynulo 96 % všech tehdejších druhů organizmů. Jak autor došel k tak vysokému číslu? Použil metodu zpětné rarefakce, která je popsána na obr. obrázek Metoda vychází ze dvou přepokladů:
- z náhodného vymírání druhů,
- z určitého, více či méně asymetrického rozložení druhů ve vyšších taxonech.
Náhodným vymíráním druhů se rozumí takový scénář, kdy jedinci jsou usmrcováni bez ohledu na to, ke kterému druhu patří; model dostal název „pěšák v poli“. Podle Raupa: Jedinci jsou vystaveni palbě a otázka života a smrti je záležitostí náhody [...] Představme si svět, ve kterém každý druh má deset jedinců, každý rod obsahuje deset druhů, každá čeleď deset rodů atd., až k jedinému kmeni s deseti třídami. Máme tedy milion jedinců [...] Jaká budou procenta vymírání pro jednotlivé taxonomické skupiny, jestliže během našeho myšlenkového experimentu zahyne sedmdesát pět procent všech jedinců? 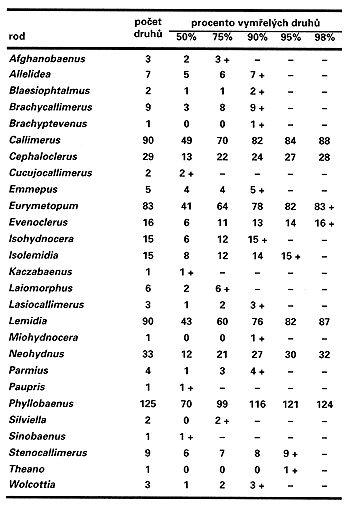 Začneme-li od nejvyššího patra, pro kmeny musí být procento vymírání nula, protože máme jen jediný kmen, jakkoli ochuzený o tři čtvrtiny příslušníků. Každá z deset tříd má sto tisíc jedinců. Existuje třída, která ztratí všechny své jedince, je-li intenzita vymírání sedmdesátipětiprocentní?
Začneme-li od nejvyššího patra, pro kmeny musí být procento vymírání nula, protože máme jen jediný kmen, jakkoli ochuzený o tři čtvrtiny příslušníků. Každá z deset tříd má sto tisíc jedinců. Existuje třída, která ztratí všechny své jedince, je-li intenzita vymírání sedmdesátipětiprocentní?  Pravděpodobnost takové události je prakticky nulová [...] Čím níže budeme v taxonomické hierarchii klesat, tím pravděpodobnější bude vymírání skupin vlivem náhody. Na samém dně tabulky je pravděpodobnost, že všech deset jedinců („pěšáků“) u kteréhokoliv druhu bude usmrceno, asi 1 : 20. Jestliže má vymírání charakter náhodného zabíjení jedinců v palbě bojiště, vyhlazením celých tří čtvrtin všech jedinců dosáhneme vymření pouhých pěti procent všech druhů. Hlavním výsledkem tohoto myšlenkového experimentu je poznání, že při náhodném zabíjení jedinců se procento vymírání týká zejména nižších stupňů taxonomického žebříčku.
Pravděpodobnost takové události je prakticky nulová [...] Čím níže budeme v taxonomické hierarchii klesat, tím pravděpodobnější bude vymírání skupin vlivem náhody. Na samém dně tabulky je pravděpodobnost, že všech deset jedinců („pěšáků“) u kteréhokoliv druhu bude usmrceno, asi 1 : 20. Jestliže má vymírání charakter náhodného zabíjení jedinců v palbě bojiště, vyhlazením celých tří čtvrtin všech jedinců dosáhneme vymření pouhých pěti procent všech druhů. Hlavním výsledkem tohoto myšlenkového experimentu je poznání, že při náhodném zabíjení jedinců se procento vymírání týká zejména nižších stupňů taxonomického žebříčku.
 Asymetrické rozložení druhů v rodech a snad i obecně nižších taxonů v taxonech vyšších je v systému organizmů známou skutečností. V zásadě platí, že (v takto studovaných systémech) většina rodů v určitém vyšším taxonu (např. čeledi nebo řádu) je monotypických nebo obsahuje jen několik málo druhů, zatímco druhově bohaté rody jsou vzácné (viz obr. obrázek) Tento předpoklad je, jak již jsem naznačil, běžně přijímán jako fakt a tvoří základ pro konstrukci křivky na obr. obrázek (v knize obr. 11, str. 64), ze které je odvozen údaj o devadesáti šesti procentech druhů vymřelých na konci permu.
Asymetrické rozložení druhů v rodech a snad i obecně nižších taxonů v taxonech vyšších je v systému organizmů známou skutečností. V zásadě platí, že (v takto studovaných systémech) většina rodů v určitém vyšším taxonu (např. čeledi nebo řádu) je monotypických nebo obsahuje jen několik málo druhů, zatímco druhově bohaté rody jsou vzácné (viz obr. obrázek) Tento předpoklad je, jak již jsem naznačil, běžně přijímán jako fakt a tvoří základ pro konstrukci křivky na obr. obrázek (v knize obr. 11, str. 64), ze které je odvozen údaj o devadesáti šesti procentech druhů vymřelých na konci permu.


 V průběhu svých studií vyšší taxonomie brouků nadčeledi Cleroidea i při pročítání entomologické taxonomické literatury jsem si všiml jedné věci: počet vyšších taxonů se snižuje v závislosti na růstu znalostí o jejich fylogenezi. Např. při úplné revizi Thanerocleridae, nové čeledi brouků, klesl počet rodů z 13 na 7, tj. téměř o 50 %. Podobně při psaní klíče rodu australských zástupců čeledi Cleridae mi klesl počet rodů z 49 na 33. V nedávné době jsem dokončil reklasifikaci Phyllobaeninae, převážně tropické podčeledi brouků čeledi Cleridae, která má optimální počet rodů a druhů pro níže popsaný experiment. Tato skupina nám bude modelem nebo, přesněji řečeno, terčem na pouťové střelnici, jak uvidíme dále. Původní počet rodů, s kterým jsem revizi začínal, byl 27 (obr. obrázek). V průběhu práce se postupně odkrývaly vztahy mezi rody i druhy, ukázalo se, že mezi mnohými druhovými skupinami, které se dosud studovaly nedostatečně a jen izolovaně, existují plynulé přechody a mnohé rody se proto ukázaly být parafyletickými (tj. určitý rod nezahrnoval všechny linie, které byly od něj odštěpeny, a ty byly klasifikovány jako samostatné rody). Takováto skutečnost se odkrývá snad před každým taxonomem, který se poctivě snaží vycházet při klasifikaci jen z toho, co vidí, používá přesné metodické postupy a studuje taxony popsané již dříve typologickými nebo evolučními taxonomy, kterým jsou podobné postupy cizí. Výsledný systém podčeledi Phyllobaeninae obsahuje po revizi jen 11 rodů, včetně jednoho nově popsaného (obr. obrázek). Počet druhů jsem pro jednoduchost ponechal podle starého systému, tj. 557 v starém i novém systému. Obrázky obrázek a obrázek ukazují rozdíly v rozložení druhů v rodech obou systémů. Je zřejmé, že křivka starého systému (obr. obrázek) se velmi podobá křivce, kterou uvádí Raup (obr. obrázek), zatímco rozložení druhů v novém systému (obr. obrázek) je mnohem vyrovnanější.
V průběhu svých studií vyšší taxonomie brouků nadčeledi Cleroidea i při pročítání entomologické taxonomické literatury jsem si všiml jedné věci: počet vyšších taxonů se snižuje v závislosti na růstu znalostí o jejich fylogenezi. Např. při úplné revizi Thanerocleridae, nové čeledi brouků, klesl počet rodů z 13 na 7, tj. téměř o 50 %. Podobně při psaní klíče rodu australských zástupců čeledi Cleridae mi klesl počet rodů z 49 na 33. V nedávné době jsem dokončil reklasifikaci Phyllobaeninae, převážně tropické podčeledi brouků čeledi Cleridae, která má optimální počet rodů a druhů pro níže popsaný experiment. Tato skupina nám bude modelem nebo, přesněji řečeno, terčem na pouťové střelnici, jak uvidíme dále. Původní počet rodů, s kterým jsem revizi začínal, byl 27 (obr. obrázek). V průběhu práce se postupně odkrývaly vztahy mezi rody i druhy, ukázalo se, že mezi mnohými druhovými skupinami, které se dosud studovaly nedostatečně a jen izolovaně, existují plynulé přechody a mnohé rody se proto ukázaly být parafyletickými (tj. určitý rod nezahrnoval všechny linie, které byly od něj odštěpeny, a ty byly klasifikovány jako samostatné rody). Takováto skutečnost se odkrývá snad před každým taxonomem, který se poctivě snaží vycházet při klasifikaci jen z toho, co vidí, používá přesné metodické postupy a studuje taxony popsané již dříve typologickými nebo evolučními taxonomy, kterým jsou podobné postupy cizí. Výsledný systém podčeledi Phyllobaeninae obsahuje po revizi jen 11 rodů, včetně jednoho nově popsaného (obr. obrázek). Počet druhů jsem pro jednoduchost ponechal podle starého systému, tj. 557 v starém i novém systému. Obrázky obrázek a obrázek ukazují rozdíly v rozložení druhů v rodech obou systémů. Je zřejmé, že křivka starého systému (obr. obrázek) se velmi podobá křivce, kterou uvádí Raup (obr. obrázek), zatímco rozložení druhů v novém systému (obr. obrázek) je mnohem vyrovnanější.
Naším úkolem je „pozabíjet“ určité procento druhů a porovnat, jak se to projeví při vymírání rodů v obou systémech. Protože nemáme po ruce nic lepšího a chceme srovnávat s Raupovými výpočty, použijeme scénář pěšáka v poli. Každý druh označíme číslem od 1 do 557. Ve starém systému budou např. druhy rodu Afghanobaenus mít čísla 1 až 3, druhy rodu Allelidea 4 – 10, ... Wolcottia 555 – 557. Generátor náhodných čísel nám vytiskl tabulku střeliva – náhodně seřazená čísla od 1 do 557, z nichž žádné se nesmí vyskytnout dvakrát (každý druh může vyhynout jen jednou). Nuže, palme! V tabulce čísel postupujme číslo po čísle, výstřel po výstřelu a z tabulek odškrtávejme zabité druhy – co číslo, to vymřelý druh. Zjistíme, že po 279 výstřelech jsme odstřelili 50 % druhů a ve starém systému nám vymřelo 15 % rodů, zatímco v novém systému žádný. Leč pokračujme ve vraždění dál! Po 418 výstřelech (75 % zabitých druhů) vymírá ve starém systému 26 % rodů, v novém pouze 18 %. 90 % vybitých druhů (501 výstřelů) je už téměř genocidou, přesto však v novém systému vymírá jen 36 % rodů. Ve starém ale téměř dvojnásobek – 63 %. Vymřely všechny rody starého systému mající do sedmi druhů, s výjimkou jediného monotypického rodu Theano, který měl v našem experimentu prostě z pekla štěstí a přežil. (Mimochodem, od jediného druhu rodu, Theano pusilla, byl znám jen jediný exemplář, holotyp, a ten je ztracen nebo alespoň dobře zašantročen kdesi v pařížském muzeu. A tak měl štěstí alespoň v naší hře.) Devadesátipětiprocenstní vymírání už se stalo osudným i rodu Theano. Ve starém systému nepřežilo 78 % rodů, kdežto v novém jen 45 %. A konečně při 98 % vyhubených druhů ještě stále přežívá celých 45 %, tj. téměř polovina rodů v novém systému, ale jen necelých 20 % rodů v systému starém.
Důvod, proč ve starém systému vymřelo více rodů, je zřejmý: obsahoval více druhově chudých rodů, které jsou při náhodném vybíjení náchylnější k vymření. V novém systému je rozložení druhů v důsledku sníženého množství rodů mnohem vyrovnanější. Vzhledem k tomu jsou ztráty na rodech při stejném množství vyhynulých druhů menší.
 Všimněme si (obr. obrázek), že nový systém déle odolává náhodnému vymírání druhů a až někde za hranicí asi 98 % dochází k prudkému úbytku rodů. Obě křivky jsou asi do osmdesáti procent vymřelých druhů přibližně stejně vzájemně vzdáleny. Spočítáme-li poměr mezi vyhynulými rody v obou systémech, dojdeme k číslu asi 0,56 (je to přibližný průměr poměrů procent vyhynulých rodů mezi padesáti a devadesáti osmi procenty vyhynulých druhů). Znamená to, že v novém systému by vymřelo téměř o polovinu rodů méně, respektive ve starém téměř dvakrát více. Raup se při svém výpočtu opírá o data založená na fosilních záznamech vymřelých vyšších taxonů mořských živočichů. Při konstrukci rarefakční křivky ovšem musel vycházet z určitého rozložení druhů mezi vyššími taxony (čeleděmi a rody). Podle průběhu samotné křivky i podle názorů obsažených v jeho knize lze usoudit, že vyšel z vysoce asymetrického rozložení. V Raupově knize se obecně popisované asymetrické rozložení druhů mezi rody (demonstrované na obr. obrázek) podobá křivce na obr. obrázek, popisující starý systém podčeledi Phyllobaeninae. Kdyby Raupova rarefakční křivka byla odvozena přímo z fosilních taxonů, pak by vycházela ze silně asymetrického rozdělení, možná ještě strmějšího než na obr. obrázek.
Všimněme si (obr. obrázek), že nový systém déle odolává náhodnému vymírání druhů a až někde za hranicí asi 98 % dochází k prudkému úbytku rodů. Obě křivky jsou asi do osmdesáti procent vymřelých druhů přibližně stejně vzájemně vzdáleny. Spočítáme-li poměr mezi vyhynulými rody v obou systémech, dojdeme k číslu asi 0,56 (je to přibližný průměr poměrů procent vyhynulých rodů mezi padesáti a devadesáti osmi procenty vyhynulých druhů). Znamená to, že v novém systému by vymřelo téměř o polovinu rodů méně, respektive ve starém téměř dvakrát více. Raup se při svém výpočtu opírá o data založená na fosilních záznamech vymřelých vyšších taxonů mořských živočichů. Při konstrukci rarefakční křivky ovšem musel vycházet z určitého rozložení druhů mezi vyššími taxony (čeleděmi a rody). Podle průběhu samotné křivky i podle názorů obsažených v jeho knize lze usoudit, že vyšel z vysoce asymetrického rozložení. V Raupově knize se obecně popisované asymetrické rozložení druhů mezi rody (demonstrované na obr. obrázek) podobá křivce na obr. obrázek, popisující starý systém podčeledi Phyllobaeninae. Kdyby Raupova rarefakční křivka byla odvozena přímo z fosilních taxonů, pak by vycházela ze silně asymetrického rozdělení, možná ještě strmějšího než na obr. obrázek.  Je to logické, protože, jak už bylo uvedeno, čím větší znalosti, tím méně vyšších taxonů – a naopak, čím menší znalosti, tím více vyšších taxonů (zejména monotypických nebo druhově chudých), a tedy asymetričtější rozdělení. Znalosti o recentních organizmech jsou jistě úplnější než o fosilních a tak systémy produkované taxonomy-paleontology jsou většinou velmi „asymetrické“. (Mimo to mám dojem, že většina paleontologů se dosud hlásí spíše k evoluční taxonomii a ke kladistice cítí bytostný odpor, což asymetřičnost jejich systémů ještě umocňuje.) Pro náš myšlenkový experiment není ovšem příliš důležité vědět, kolik parafyletických skupin Raupův modelový systém obsahuje (to ani vědět nemůžeme), jde nám jen o to ukázat, jak změna taxonomického přístupu ke klasifikaci pohne s výsledkem při rarefakční metodě odhadu vyhynulých druhů.
Je to logické, protože, jak už bylo uvedeno, čím větší znalosti, tím méně vyšších taxonů – a naopak, čím menší znalosti, tím více vyšších taxonů (zejména monotypických nebo druhově chudých), a tedy asymetričtější rozdělení. Znalosti o recentních organizmech jsou jistě úplnější než o fosilních a tak systémy produkované taxonomy-paleontology jsou většinou velmi „asymetrické“. (Mimo to mám dojem, že většina paleontologů se dosud hlásí spíše k evoluční taxonomii a ke kladistice cítí bytostný odpor, což asymetřičnost jejich systémů ještě umocňuje.) Pro náš myšlenkový experiment není ovšem příliš důležité vědět, kolik parafyletických skupin Raupův modelový systém obsahuje (to ani vědět nemůžeme), jde nám jen o to ukázat, jak změna taxonomického přístupu ke klasifikaci pohne s výsledkem při rarefakční metodě odhadu vyhynulých druhů.
 Raupových 96 % vyhynulých druhů v permské katastrofě odpovídá na jeho rarefakční křivce osmdesáti šesti procentům vyhynulých rodů. Když vynásobíme těchto 86 % naším koeficientem 0,56, dostaneme číslo 48. Předpokládáme tedy, že pokud by Raupův systém prošel důkladnou taxonomickou revizí (při které by ovšem byla k dispozici úplnější data, než bývají u fosilního materiálu), pak by počet vymřelých rodů byl téměř poloviční, tj. 48 % oproti Raupovým 86 %. Nyní můžeme použít metodu zpětné rarefakce k novému odhadu počtu vymřelých druhů při permské katastrofě. Vycházejme i nadále z předpokladu, že Raupův systém by se po revizi změnil podobně, jako se změnil systém podčeledi Phyllobaeninae. Rozdílů mezi křivkami pro oba systémy podčeledi Phyllobaeninae proto použijeme pro sestrojení nové rarefakční křivky pro Raupův systém „po revizi“ (obr. obrázek). 48 % vymřelých rodů na nové křivce odpovídá osmdesáti sedmi procentům vyhynulých druhů. Vzhledem k předpokládanému symetričtějšímu rozložení druhů v rodech (rody jsou nyní druhově bohatší) poklesl tak počet vymřelých rodů o 38 % a počet vymřelých druhů o 9 %. A to jen změnou taxonomické metodiky při klasifikaci organizmů, které byly použity v rarefakční metodě odhadu, tedy vlastně jen pouhým administrativním opatřením. A to je prosím výsledek našeho myšlenkového experimentu nebo ukázky, co všechno může ovlivnit výpočty tohoto typu.
Raupových 96 % vyhynulých druhů v permské katastrofě odpovídá na jeho rarefakční křivce osmdesáti šesti procentům vyhynulých rodů. Když vynásobíme těchto 86 % naším koeficientem 0,56, dostaneme číslo 48. Předpokládáme tedy, že pokud by Raupův systém prošel důkladnou taxonomickou revizí (při které by ovšem byla k dispozici úplnější data, než bývají u fosilního materiálu), pak by počet vymřelých rodů byl téměř poloviční, tj. 48 % oproti Raupovým 86 %. Nyní můžeme použít metodu zpětné rarefakce k novému odhadu počtu vymřelých druhů při permské katastrofě. Vycházejme i nadále z předpokladu, že Raupův systém by se po revizi změnil podobně, jako se změnil systém podčeledi Phyllobaeninae. Rozdílů mezi křivkami pro oba systémy podčeledi Phyllobaeninae proto použijeme pro sestrojení nové rarefakční křivky pro Raupův systém „po revizi“ (obr. obrázek). 48 % vymřelých rodů na nové křivce odpovídá osmdesáti sedmi procentům vyhynulých druhů. Vzhledem k předpokládanému symetričtějšímu rozložení druhů v rodech (rody jsou nyní druhově bohatší) poklesl tak počet vymřelých rodů o 38 % a počet vymřelých druhů o 9 %. A to jen změnou taxonomické metodiky při klasifikaci organizmů, které byly použity v rarefakční metodě odhadu, tedy vlastně jen pouhým administrativním opatřením. A to je prosím výsledek našeho myšlenkového experimentu nebo ukázky, co všechno může ovlivnit výpočty tohoto typu.
V žádném případě se nechci paleontologům vnucovat se svými osmdesáti sedmi procenty druhů vyhynulých koncem permu, ačkoli můj postup snad nebyl o nic méně regulérní než Raupův. Ovšem scénář pěšáka v poli není při katastrofách univerzálně platný, jak ukazuje např. selektivní vyhynutí trilobitů koncem permu nebo dinosaurů v křídě. „Špatné geny“, ať už to znamená cokoli, jistě existují. Kromě toho při permské katastrofě se většinou udává nepoměr mezi vymíráním ve vodě a na souši (suchozemské organizmy byly postiženy méně) a Raupovy odhady jsou založeny na mořských živočiších.
Literatura
D. M. Ruap: O zániku druhů. NLN, Praha 1995P. R. Erlich, D. D. Murphy: Butterflies and biospecies. J. Res. Lepid. 21, 219 – 225, 1983
L. A. Mound: Biological reality – the taxonomists` chimaera. Atti XIV Congr. naz. ital. Ent. Palermo, 135 – 147, 1985
J. Kolibáč: Revision of Thanerocleridae n. stat. (Coleoptera, Cleroidea). Mitt. schweiz. ent. Ges. 65, 303 – 340, 1992
DEFINICE DRUHU A VYŠŠÍCH TAXONŮ
Obecně lze říci, že je tolik definic druhu, kolik je přírodovědců, a to včetně tvrzení relacionistů, že žádné druhy jako objektivní jednotky v přírodě neexistují.Učebnicová definice biologického bisexuálního druhu zní takto: „Druh je skupina aktuálně nebo potenciálně se křížících populací, které jsou reprodukčně izolovány od jiných takových skupin“ (podle E. Mayra). Klasická Mayrova definice rozeznává dvojí izolační mechanizmy: předkopulační a pokopulační. Do první skupiny patří např. rozdílná ekologie nebo etologie jednotlivých druhů, do druhé např. neživotnost zárodků kříženců, neplodnost kříženců atd. Alternativou zmíněné definice je MRC (mate-recognition species concept), kterou propaguje skupina jihoafrických biologů. Ti tvrdí, že druhy jsou „samodefinující“, a proto vyloučili ze své definice pokopulační izolační mechanizmy. Druh je podle nich „uzavřená skupina populací bisexuálních organizmů, které sdílejí společný rozmnožovací systém.“
Mimo to ještě existuje řada definic „taxonomického“ druhu, které v podstatě říkají, že druh je soubor jedinců sdílejících společné znaky. Tyto definice jsou nejjednodušší a také mezi taxonomy nejpoužívanější. Taxonomové, kteří jsou nuceni pracovat převážně s mrtvými muzejními exempláři (např. mnozí entomologové), většinou nemají znalosti izolačních mechanizmů a jejich metody musí být ve shodě s pravidly zoologické nomenklatury. Ačkoli si to zřídka uvědomují, jejich druhový koncept bývá křížencem mezi starým linnéovským typologickým pojetím a MRC. Většina živočišných druhů je tedy popisována na základě následujícího konceptu: druh je skupina jedinců, kteří mají společný rozmnožovací systém s holotypem. Recept na klasifikaci živočišných druhů by podle toho zněl asi takto: (1) ze skupiny živočichů vyber libovolného jedince a označ jej jako holotyp; (2) ze zbytku vyber jedince s natolik společnými znaky, že u nich předpokládáš společný rozmnožovací systém s holotypem; (3) jestliže zbývají nezařazení jedinci, jdi na (1); (4) konec. To je základní algoritmus tzv. alfa-taxonomie. A tak druh je prostě to, co taxonom jako druh označí.
Nakonec ještě Willeyho „evoluční druhový koncept“, který osobně považuji pro jeho dynamičnost za nejbližší skutečnosti: „Evoluční druh je jediná linie populací od předka k potomkům, která se odlišuje od jiných takových linií a má své vlastní evoluční tendence a historii.“ (Mimochodem, jak pevný asi musí být chrám biologické vědy a naopak jak složitá a vědecky těžko pochopitelná je příroda, když nejsme schopni přesně definovat ani tu nejzákladnější jednotku biologie – to jest druh.)
Vyšší taxony (VT) jsou soubory druhů (u rodů, podrodů) nebo nižších VT (u čeledí, řádů, tříd, kmenů, říší a mezistupňů) založené na předpokládané příbuznosti. VT tedy jsou taxonomické kategorie, které by měly vyjadřovat fylogenetické vztahy mezi organizmy. V zásadě existují dva hlavní přístupy k vyšší taxonomii. První, evolucionistický, praví, že VT jsou skupiny nějak odlišné od ostatních skupin stejné kategorie. Tento přístup umožňuje tzv. negativní definování VT. Např. na oddělení rodu A od rodu B postačí udat, že A má znak, který B chybí. Druhý, kladistický přístup negativní definování neumožňuje a vyžaduje od taxonoma, aby všechny VT byly založeny na evolučně pokročilých znacích (apomorfiích). Takové VT se nazývají monofyletické. Jaká konkrétní taxonomická kategorie (např. zda tribus nebo podčeleď) bude monofyletickému VT přidělena, ovšem do značné míry závisí na subjektivním rozhodnutí taxonoma. VT jsou proto v současné taxonomii do značné míry arbitrárními konstrukcemi. Osobně jsem ale názoru, že VT jsou v přírodě objektivně se vyskytujícími entitami právě tak jako druh a lze je definovat podobně jako ve výše zmíněném evolučním druhovém konceptu.
O autorovi
Jiří Kolibáč

Doporučujeme

Ničí ozon choleru? 

Jak se člověk stává biologem
